
Ecological Theory and the Superfluous Niche
James Justus
Philosophical Topics, Volume 47, Number 1, Spring 2019, pp. 105-123 (Article)
Published by University of Arkansas Press
For additional information about this article
[ Access provided at 26 Nov 2020 14:45 GMT from The University of Guelph ]
https://muse.jhu.edu/article/774175
105
. , . ,
Ecological eory and the Superfluous Niche
James Justus
Florida State University
ABSTRACT: Perhaps no concept has been thought more important to
ecological theorizing than the niche. Without it, technically sophisticated
and well-regarded accounts of character displacement, ecological equiva-
lence, limiting similarity, and others would seemingly never have been
developed. e niche is also widely considered the centerpiece of the
best candidate for a distinctively ecological law, the competitive exclu-
sion principle. But the incongruous array and imprecise character of pro-
posed denitions of the concept square poorly with its apparent scientic
centrality. I argue this denitional diversity and imprecision reects a
problematic conceptual indeterminacy that challenges its putative indis-
pensability in ecology.
Perhaps no concept has been thought more important to ecological theorizing
than the niche. Without it, technically sophisticated and well- regarded accounts
of character displacement, ecological equivalence, limiting similarity, and others
would seemingly never have been developed. e niche is also widely considered
the centerpiece of the best candidate for a distinctively ecological law, the competi-
tive exclusion principle. e received view in ecology has therefore been that the
niche is indispensable, despite occasionally vocal protests from a small minority.
Aer all, the concept is oen said to simply precisify the idea that species make
their biological livelihoods in dierent ways, and what could be more central to
ecology? Many (if not most) inuential analyses in the 1960s and 1970s bore the
106
‘niche’ label, oen paying homage to Hutchinson’s (1957) highly abstract denition
in particular. Mechanistic models of resource consumption that predominated
subsequent decades (e.g., Tilman 1982) are taken to extend and rene the same
approach, the niche similarly at their core. More recently, Hubbell’s (2001) unied
neutral theory certainly perturbed the prevailing assessment, but it fell far short
of upending it (see Odenbaugh forthcoming). A prominent book responding to
neutralist theories in favor of niche- based theorizing, for instance, proclaims “the
niche has provided and can continue to provide the central conceptual foundation
for ecological studies” (Chase and Leibold 2003, 17).
In this case, however, the naysayers were right. e incongruous array and
imprecise character of proposed denitions of the concept square poorly with its
apparent scientic centrality. Rather than reect innocuous semantic dierences
or a potentially useful integrative pluralism, this denitional diversity and impre-
cision reects a problematic conceptual indeterminacy that challenges its putative
indispensability in ecology. e niche has not and cannot— at least as it has been
characterized thus far— do the substantive, foundational work it is claimed to do
in ecology. e conceptual content tethered to the term ‘niche’ is just too problem-
atically disjoint and amorphous to play that role.
e gap between conceptual aspiration and scientic practice permeates apprais-
als of ecological theorizing to the present in many dierent, multifaceted ways. is
analysis focuses specically on the concept’s origins in the work leading up to, and
in many ways culminating in, Hutchinson’s highly abstract n- dimensional hyper-
volume denition. Section 1 describes the emergence of the ecological niche in
Joseph Grinnell and Charles Elton’s work. From the very beginning, the concept’s
content was unmistakably disjoint: environments and how they impact species was
one focus; how species function in communities, particularly via trophic inter-
actions, was the other. Beyond the bivalent focus, the concept was also problem-
atically imprecise. is point is illustrated by considering the contentious idea of
“vacant” niches and the signicant indeterminacy about their possibility.
Despite a widespread view it is, section 2 argues the niche concept is not the
centerpiece of perhaps the best candidate for a distinctively ecological law, the com-
petitive exclusion principle. Gause’s (1934) Paramecium experiments and putatively
mechanistic explanation of competitive dynamics with Lotka- Volterra equations
are widely taken to supply the rst compelling grounds for a niche- based version
of the exclusion principle (see Hutchinson 1978). But the evidence for this judg-
ment is pretty thin. Gause’s explanation in e Struggle for Existence actually makes
little use of niche ideas. His semantically suggestive— but thoroughly non- niche—
term ‘vacant places’ may have led many later commentators astray. And what little
he did say about niches provides little guidance about how niche considerations
might be brought to bear on models of competitive dynamics, via competition
coecients for instance. is serious deciency is a quite general problem, one
shared by later attempts to characterize the concept, including its most inuential
denition in ecology.

107
at denition is Hutchinson’s n- dimensional hypervolume characterization
of the niche, the focus of section 3. As judged by the attention it received, his de-
nition had an enormous impact on ecology. But judged by the content conveyed,
the impact seems disproportionate. Hutchinson made highly questionable and
signicantly limiting assumptions in characterizing the concept. e most seri-
ous deciency, however, is the same kind of conceptual impoverishment exhibited
by earlier attempts to pinpoint the concept. Section 4 concludes by arguing that
Chase and Leibold’s (2003) more recent revival of the niche is problematic in the
same way. Rather than convey information about community dynamics, infor-
mation that helps represent and analyze those dynamics, the niche superuously
supervenes on them on their account of the concept.
1. GRINNELL AND ELTON’S NICHES
e rst director of the Museum of Vertebrate Zoology at the University of
California at Berkeley, Joseph Grinnell, wasn’t the rst to use ‘niche’ in an ecologi-
cal sense,
1
but he was the rst to do so with any signicance. His most well- known
paper doing so— the rst ecological publication with ‘niche’ in the title— contains
three instances all in the penultimate paragraph:
ese various circumstances, which emphasize dependence upon cover,
and adaptation in physical structure and temperament thereto, go to
demonstrate the nature of the ultimate associational niche occupied
by the California rasher. is is one of the minor niches which with
their occupants all together make up the chaparral association. It is, of
course, axiomatic that no two species regularly established in a single
fauna have precisely the same niche relationships. (Grinnell 1917, 433)
2
e “circumstances” are the chaparral habitat’s physical characteristics, for which the
rasher’s phenotypic properties are especially well suited. Its dense undercanopy
foliage prevents all but short bursts of ight, complementing the rasher’s small,
compact wings. Its inconspicuous drab- brown plumage also enhances predator-
evasion in that foliage.
e allusions to occupation are important. Grinnell’s primary focus was ani-
mal species; the vegetation they inhabit was conceptualized as part of their physical
environments. Niches are then units of that physical, partly biotic environment for
Grinnell, units species can occupy. With evolutionary history in mind, relationships
between occupants and what’s occupied can therefore be explanatory: properties of
1. Apparently, Roswell Johnson was in a 1910 study of ladybug color patterns (see Hutchinson 1978,
155–56).
2. Note the last sentence’s close similarity with what was later labeled the ‘competitive exclusion
principle’ (see §2).

108
niches, as actual bits of physical space, can account for why organisms residing in
them possess the (adaptive) phenotypic properties they do.
Although a minority view, there are contrasting readings of Grinnell. Hutchinson
(1978, 157) claimed, “it is evident that [for Grinnell] the space occupied by ‘just
one species’ is an abstract space that cannot be a subdivision of the ordinary habi-
tat space.” But such abstraction coheres poorly with Grinnell’s extensive descriptions
of actual portions of environments as niches. e emphasis on the actual is quite
clear in later papers:
Habitats have been variously classied by students of geographical dis-
tribution. Some of us have concluded that we can usefully recognize, as
measures of distributional behavior, the realm, the region, the life- zone,
the fauna, the subfauna, the association, and the ecologic or environ-
mental niche. e latter, ultimate unit, is occupied by just one species or
subspecies; if a new ecologic niche arises, or if a niche is vacated, nature
hastens to supply an occupant, from whatever material may be available.
Nature abhors a vacuum in the animate world as well as in the inanimate
world. (Grinnell 1924, 227)
Note that habitats are being classied with dierent measures of geographical dis-
tribution, the unit of nest resolution being the “ecologic or environmental” niche
(synonymy implied).
3
e idea an abstraction is really what’s being invoked there-
fore appears implausible. at Hutchinson favored and developed an abstract- space
approach himself may be relevant.
In one of ecology’s founding works, Animal Ecology, Charles Elton (1927, 63–64)
described a very dierent concept:
Animals have all manner of external factors acting upon them— chemical,
physical, and biotic— and the “niche” of an animal means its place in the
biotic environment, its relations to food and enemies. e ecologist should
cultivate the habit of looking at animals from this point of view as well as
from the ordinary standpoints of appearance, names, anities, and past
history. When an ecologist says “there goes a badger” he should include
in his thoughts some denite idea of the animal’s place in the community
to which it belongs, just as if he had said “there goes the vicar.”
Elton recognized the impacts of abiotic (“chemical,” “physical”) factors, but unlike
Grinnell his niche focuses on biotic interactions. e one diagram presented in
the ‘Niches’ section, for example, illustrates the “niche occupied by small sap-
suckers, of which one of the biggest groups is the plant- lice or aphids” (1927,
66) with a ‘food- cycle’— ‘food- web’ in current terminology— comprised solely of
biological nodes:
3. ‘Fauna’, ‘subfauna’, and ‘association’ were not categories of biological composition for Grinnell.
Rather, they were hierarchical units of geography— determined primarily by abiotic factors such
as humidity and temperature— within which species distributions could be classied and hypothe-
sis about their causes evaluated (Griesemer 1990).
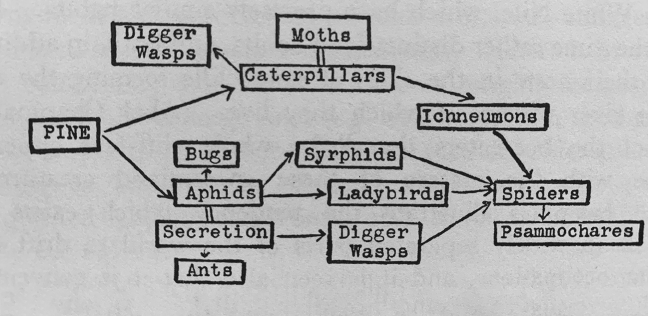
109
Figure 1. Food-cycle on young pine-trees on Oxshott Common.
is gure and reference to a “small sapsuckers” niche (note the plural) reveals
an interesting aspect of Elton’s concept absent from Grinnell’s. For Elton, what
individuates niches partially depends on how ne- grained the relations between
organisms comprising a biological community are conceptualized. On this point
Elton (1927, 64) was quite explicit:
[W]e might take as a niche all the carnivores which prey upon small
mammals, and distinguish them from those which prey upon insects.
When we do this it is immediately seen that the niches about which
we have been speaking are only smaller subdivisions of the old con-
ceptions of carnivore, herbivore, insectivore, etc., and that we are only
attempting to give more accurate and detailed denitions of the food
habits of animals.
Degree of representational resolution therefore determines what counts as a niche.
And since that degree diers across scientic contexts, species are constituents of
many distinct niches with dierent extensions. e claim that coexisting species
cannot have the same niche— one gloss on the competitive exclusion principle that
Grinnell expressed in the rst quote above— would thus sound foreign to Elton. It
relies on a much narrower niche conception. is greater generality may account for
the fact, noted by Hutchinson (1978, 152), that Elton never used the niche concept
to explain competition. It likely also explains why he was never tempted to elevate
competitive exclusion to axiomatic or principle status.
Dependence on representational resolution therefore marks an important con-
trast between Grinnell and Elton’s understandings of niche. But the contrast also
seems to stem from a much deeper and more signicant divergence of habitat vs.
functional conceptions. Unlike Grinnell’s markedly physical conception, Elton’s
vicar analogy manifests the latter. Just as vicars are identied by functional roles in

110
religious institutions, an animal’s “place in the community” is similarly function-
ally individuated by its role in networks of biotic interactions.
Some resist this bipartite judgment (e.g., Griesemer 1992; Schoener 1989).
Griesemer rightly stresses that Grinnell and Elton had dierent research foci—
primarily, how evolutionary dynamics inuence species distributions vs. how
trophic interactions determine community structure— and that that probably par-
tially explains their divergent uses of “niche.” He also correctly highlights that each
recognized the importance of biotic and abiotic factors, while obviously believing
one was more salient than the other. e claim Elton and Grinnell possessed dis-
tinct biotic and abiotic concepts should therefore be rejected. ese dierences,
though nontrivial, only indicate dierent conceptual emphases for Griesemer, not
dierent concepts: “Grinnell and Elton both identied the niche as the place/role
a species happens to occupy in an environment” (1992, 235).
Correctly judging when dierences in conceptual emphasis signify distinct
concepts requires a theory of concept individuation, which is beyond the pur-
view of Griesemer’s analysis (or this one). But some of the dissimilarities seem
to make concept- individuating dierences. For instance, the clause ‘occupy in an
environment’ in Griesemer’s characterization is problematic in Elton’s case. Elton
did mention an animal’s place in the ‘biotic environment’— though just once in
Animal Ecology (see above)— but the immediate, emphasized paraphrase (“its rela-
tions to food and enemies”) strongly suggests the functional sense given explicitly
two sentences later in the vicar analogy with the very similar phrase “animal’s place
in the community.”
Elton’s recognition of a small- mammal- consuming- carnivore niche and car-
nivore niche in toto reinforces this conclusion. ese classes are not characterized
in relation to environments their members occupy, but rather by their functional
role in communities: consuming eshy prey. On this issue Elton was unambigu-
ous. e ‘Niches’ section begins, “[A]lthough the actual species of animals are
dierent in dierent habitats, the ground plan of every animal community is
much the same. In every community we should nd herbivorous and carnivo-
rous and scavenging animals.” Aer giving specic examples the paragraph con-
tinues, “It is therefore convenient to have some term to describe the status of an
animal in its community, to indicate what it is doing and not merely what it looks
like, and the term used is ‘niche’ ” (1927, 63). Niches are thereby characterized
in terms of this common ‘ground plan’: an abstract pattern of basic functional
relations underlying all communities according to Elton. But that plan is con-
trasted with the dierent habitats species inhabit. Irrespective of whether niches
are ‘places’ or ‘roles’— two terms that oen have very dierent connotations— for
Elton they are not parts of environments. is concept, unlike Grinnell’s, is thor-
oughly functional.
4
4. Elton himself sharply distinguished his concept from Grinnell’s. He criticized Odum’s Fundamentals
of Ecology for failing to distinguish the two (Elton 1954).

111
ere is, however, a more general perspective from which this dierence can
appear artifactual. Both Grinnell and Elton describe niches as components of
broader patterns, be they structures in physical environments or networks of func-
tional interactions. If these patterns derive from or simply are causal relations, then
both ecologists are giving causal representations, the only dierence being the nature
of the causal relata. is dierence would then just reect Grinnell and Elton’s dier-
ent investigative priorities and explanatory commitments; the underlying content of
the niche concept would be the same. e unied characterization would then be:
Niche— a node in a nexus of causal interactions with abiotic and biotic
factors occupied by a species.
In a ne- grained species- specic way, ‘niche’ would then simply convey causal
information about ecological systems.
Despite the theoretical allure of unication, this proposal clearly fails. e
problem is that Grinnell and Elton both countenanced the possibility of empty or
vacant niches: unoccupied parts of environments (see Grinnell 1924, 227, quoted
above), or biologically uninstantiated constituents of food webs (see Elton 1927,
27). e causally focused, species- specic characterization above seems unable
to capture this broader notion; tracking the causal habits of nonexistent species is
quite dicult. And this wasn’t a superuous conceptual aside. e idea was thought
to do important work. Vacant niches feature prominently in Grinnell’s explanations
of the adaptiveness of species’ phenotypes for specic environments, and Elton’s
explanations of putative ecological equivalents in distinct biological communities.
5
One might think there is an easy x, simply add “or not” to the characteriza-
tion above. But the vacant disjunct is not so conceptually innocuous. Its addition
seems to abandon the very causal information upon which the unied charac-
terization is based. If it is the web of causal relations a species realizes that indi-
cates the contours of the niche it occupies, no indication occurs in empty cases
(from nonexistent species). Yet Grinnell and Elton were committed to the idea that
niches endure rather than expire when those causal relations cease to exist (niches
are vacated). So they presumably cannot be (part of?) what constitutes a niche.
But what, then, is the relationship between the causal relations species partici-
pate in when they occupy a niche and whatever it is that characterizes that niche?
Far from being “formalized by Grinnell” (Chase and Leibold 2003, 8), without an
answer to this question the niche concept in its Grinnellian, Eltonian, or causal-
unicatory guises is problematically vague.
Vagueness is not always problematic in science. Imprecision can accurately
represent appropriate uncertainty about how a phenomenon is best described, or
capture precisely the right level of generality when explaining it. It is problematic
5. A more recent example is Lawton’s (1982) inuential analysis of bracken herbivores in North
American and Britain, and conclusion that the American communities were comparatively
“unsaturated”— containing many more empty niches— and were therefore more susceptible to
invasion.
112
here because vagueness precludes clarity about what individuates niches. And
clear individuation standards are certainly necessary if the concept is to perform
the substantive function it is thought to in ecological theorizing. is kind of criti-
cism has a well- established track record in biology. It is the same kind of charge
made against adaptationism in an evolutionary context:
e niche is a multidimensional description of all the relations entered
into by an organism with the surrounding world... To maintain that
organisms adapt to the environment is to maintain that such ecological
niches exist in the absence of organisms and that evolution consists
in lling these empty and preexistent niches. But the external world
can be divided up in an uncountable innity of ways, so there is an
uncountable innity of conceivable ecological niches. Unless there is
a preferred or correct way in which to partition the world, the idea of
an ecological niche without an organism lling it loses all meaning.
(Levins and Lewontin 1985, 68)
Allusion to uncountable innities aside, it certainly doesn’t seem that Grinnell or
Elton’s accounts provide much guidance about how to aect the required partitioning.
For the kind of purpose Grinnell and Elton oen seemed to have in mind,
however, the inability to partition might be only marginally problematic, if at all.
Rather than attempting to ecologically carve nature at some joint— one strictly
inhabitable by a single species— they were oen concerned with analogical infer-
ences across dynamically and structurally similar communities. Communities
in geographically and evolutionarily remote areas oen seem to exhibit similar
dynamics. If this African grassland community has a large species guild perform-
ing this critical ecosystem function, then that seemingly analogous American
grassland community likely has a similarly large guild performing that same kind
of function. is type of inference is particularly clear in Elton’s allusion to a basic
‘ground plan’ underlying all animal communities. But the same idea underlies
Grinnell’s descriptions of how species are adapted to properties of their physical
environments, and how at very distant geographical locations one still oen nds
similar kinds of species if their environments are similar. Lawton’s (1982) bracken
study is another example of the same kind of inference.
ese analogical inferences depend on recognizing broad causal patterns that
indicate similarities of structure or dynamics: the similarities make the analogies
apt. But analogy aptness doesn’t require isomorphism, homomorphism, or any
other ne- grained correspondence of dynamics or structure, the kind of precise
correspondence a niche concept that partitioned the environment (Grinnell) or
functional community relations (Elton) would aord. Of course, analogical infer-
ences are notoriously dicult to assess. But when they are fruitful it’s charac-
teristically not because such a high degree of correspondence precision can be
established. Such precision in fact cuts against the less constrained connections
analogies trade upon. Biologically informed pattern recognition, not a founda-
tional and systematic niche concept, seems to underpin the cross- community,
cross- environment inferences Elton and Grinnell were making.

113
2. THE STRUGGLE FOR EXISTENCE AND
COMPETITIVE EXCLUSION
e shortcoming discussed above squares poorly with the prevalent view of the
niche’s role in perhaps the best candidate for a distinctively ecological law, the
competitive exclusion principle (CEP). Simply put, it says complete competitors
cannot coexist (Hardin 1960) or, in niche- theoretic terms, species with identical
niches cannot coexist. is principle has a long history in ecology. Grinnell, for
instance, arrived at an exclusionary principle early in the twentieth century, per-
haps drawing upon suggestive passages from Darwin (see Hardin 1960).
6
But the
CEP’s most compelling development and elevation to ‘principle’ status is largely
credited to Georgii Gause, his inuential e Struggle for Existence in particular.
Despite its pedigree and pretensions to lawhood, the CEP is controversial.
With Popperian are, Peters (1991) deemed it tautologous. Even more methodo-
logically tolerant ecologists have called it “untestable” and “of little scientic util-
ity” (Pianka 2000, 248). e present task isn’t to render judgment on these claims.
Rather, it’s to evaluate what could be considered the received view about the con-
tribution the niche concept makes to CEP, which Griesemer (1992, 237) captures:
“Gause’s and Park’s experiments showed that the concept of niche, in the guise of
determinants of relations of competitive exclusion, was central to an understand-
ing of population dynamics and the evolutionary structuring of communities.”
But if the concept is as problematically imprecise as indicated in section 1, such
centrality would be perplexing. No concept that indeterminate can do that much
heavy theoretical liing. Fortunately, a close reading of Gause’s reasoning shows
that although he used the term, the concept actually contributes little.
In several ingenious experiments Gause (1934) studied competitive dynamics
in Paramecium and yeast species. In constant ecological conditions (e.g., nutrient
levels, medium temperature, turbidity, etc.), and absent refugia that might miti gate
interspecic competition eects, one species inevitably outcompeted the other to
extinction. is result matches what classical Lotka- Volterra competition equations
predict (exclusion), and Gause believed this furnished a compelling case for CEP.
e key question is what the niche concept contributes to this case.
Gause (1934, 19) rst mentioned ‘niche’ in a context- setting discussion of
“general principles” zoologists had developed in connection with competition.
Aer citing Elton’s (1927) “place in a community” denition referencing “habits,
food, and mode of life,” Gause then stated, “It is admitted that as a result of com-
petition two similar species scarcely ever occupy similar niches, but displace each
other in such a manner that each takes possession of certain peculiar kinds of food
and modes of life in which it has an advantage over its competitor.”
7
e clause “It
6. “[T]wo species of approximately the same food habits are not likely to remain long evenly bal-
anced in numbers in the same region. One will crowd out the other” (Grinnell 1904, 377).
7. Gause (1934) did not cite and was apparently unaware of Grinnell’s work.
114
is admitted” reects Gause’s awareness that Lotka, Volterra, and J. B. S. Haldane
(1924) had already demonstrated exclusion with mathematical models of compe-
tition, models in which ‘niche’ is absent (see below). e CEP was denitely “in the
air” well before e Struggle.
But what is most striking about Gause’s claim is how little Elton (1927) actu-
ally tied the niche concept to competition, and that he didn’t entertain anything
resembling the CEP. In fact, Elton allowed that two species might occupy one niche
(see §1). While many ecologists at the time were seeing competition as the prime
driver of community structure (Kingsland 1995), Elton never shared this con-
dence. Elton was also quite skeptical of the ecological salience of mathemati cal
approaches to studying natural systems; he thought they typically over simplied
their subject (Crowcro 1991). Gause’s eort to situate his project within the inu-
ential work of the day— Animal Ecology having had an immense impact on the
incipient science— therefore seems to run a bit roughshod over the actual content
of Elton’s niche concept.
Immediately aer mentioning Elton’s niche, Gause (1927, 19–20) illustrated
the idea of closely related species with dierent niches with an example of phylo-
genetically close sympatric tern species. ey appeared to minimize competition
by having distinct food sources. But the importance of dierential feeding behav-
iors on communities composed of related species was well known long before
Elton (or Grinnell) ecologically coined ‘niche,’ at least since Darwin’s discussion
of Galapagos nches. Moreover, modeling work bere of niche considerations
Gause knew well demonstrated the same result. Gause (1934) cited Lotka’s (1932)
analysis of competitive equations, which showed competitors could coexist by uti-
lizing dierent food sources. e analysis never mentioned ‘niche’ and Elton and
Grinnell are not referenced. Without something beyond the mere fact that Elton’s
niche includes food, it therefore appears the concept contributes little in this part
of e Struggle to Gause’s case for CEP.
e aim of Gause’s initial discussion was context- setting, however. And at the
section’s end Gause (1934, 19) emphasized, “we shall endeavor to express all these
relations [food sources on competition] in a quantitative form.” His explanation of
how Lotka- Volterra dierential equations represent competitive dynamics 25 pages
later supplies that quantication, and it contains the next ‘niche’ reference. If the
niche concept is to make a signicant contribution to the CEP, this is the place.
At rst glance, the intended contribution seems clear. ‘Niche’ rst occurs in
this section in Gause’s (1934, 45–46) discussion of α, a coecient of interspecic
competition in the equations:
if the interests of the dierent species do not clash and if in the micro-
cosm they occupy places of a dierent type or dierent “niches” then
the degree of inuence of one species on the opportunity for growth
of another, or the coecient α, will be equal to zero. But if the species
lay claim to the very same “niche,” and are more or less equivalent
as concerns the utilization of the medium, then the coecient α will
approach unity.

115
Putting the potential signicance of scare quotes aside, niches— via ‘places’— then
seem to factor explicitly into Gause’s (1934, 47) word- equation- explanation of the
equations given one page later:
Immediately aer this verbal description, Gause (1934, 47) gave their mathemati-
cal representation:
(1a)
(1b)
where N
1,2
represent competing species; b
1,2
represent birth rates; K
1,2
represent
“maximally possible” carrying capacity population sizes; and α, β are competition
coecients representing the eect of N
1
individuals on N
2
individuals, and vice
versa. ese equations for the “struggle for existence,” Gause (1934, 48) claried,
“express quantitatively the process of competition between two species for the pos-
session of a certain common place in the microcosm.” Note ‘place’ here and in the
word- equation.
“Degree of realization of the potential increase” in the far right term desig-
nates the “drag” factors represented in competition equations. (1) shows there are
two: intraspecic density- dependent drag captured by the logistic element (the
second term: ) and interspecic density- dependent drag captured
by the competition element (the third term: ). ese factors,
the word- equation tells us, depend upon the “number of still vacant places.” What
else could those places be but the same “places” Gause considered when discussing
α and “niches” a single page before, or the “place” in Elton’s niche characteriza-
tion Gause referred to explicitly? e rst chapter of e Struggle pays Darwin

116
signicant tribute for largely founding its scientic subject matter; perhaps Gause
was harkening back to his niche- like use of ‘place’ (see Pearce 2010).
8
e terminological convergence here, however, is highly misleading. By ‘place’
in “vacant places” and “common place in the microcosm” Gause meant something
very specic, and entirely distinct from ‘place’ in Elton’s “place in a community.”
e former sense is made clear in Gause’s earlier discussion of the logistic equa-
tion’s representation of intraspecic density- dependence. For species N in a spe-
cic environment at a particular time, “e dierence between the maximally
possible and the already accumulated population (K – N), taken in a relative form,
i.e., divided by the maximal population , shows the relative number of ‘still
vacant places’ ” (Gause 1934, 34–35). at is, ‘vacant places’ simply numerically
measure how much more populations can grow given intraspecic density-
dependence and— when competing with another species— interspecic density-
dependence. ‘Common places’ are then actually arithmetic units of population
size, realized or potential (vacant), that species “compete” for according to Gause.
9
Function- laden Eltonian notions of habit, feeding behavior, and mode of life are
orthogonal to this numeric ‘place’ notion. e former therefore doesn’t help make
the case for CEP via the latter.
But this portion of e Struggle— the mathematically rich explanation of
competition— is arguably its most compelling core: “Gause’s great achievement
was to give a clear exposition of the way that competitive exclusion, so oen previ-
ously noted, actually worked” (Hutchinson 1978, 152). e ostensibly mechanistic
detail of the account of competitive dynamics made it so convincing. If niche-
considerations contribute little if anything to that account, what real work is the
concept doing in Gause’s case for CEP?
e remaining ve ‘niche’ references in e Struggle bolster this judgment.
ey occur (with occasional scare quotes again) 50 pages later in Gause’s discus-
sion of several Paramecium experiments that resulted in competitive exclusion.
All those references appear in one passage questioning the result’s relevance to
real- world biological systems:
However, there is in nature a great diversity of “niches” with dierent
conditions, and in one niche the rst competitor possessing advantages
over the second will displace him, but in another niche with dierent
conditions the advantages will belong to the second species which will
completely displace the rst. erefore side by side in one community,
but occupying somewhat dierent niches, two or more nearly related
8. In fact, adding to the terminological congruence, Gause (1934) seemingly used it in precisely
this sense on page 1: “Darwin considered the struggle for existence in a wide sense, including the
competition of organisms for a possession of common places in nature, as well as their destruc-
tion of one another.”
9. is language is very strained. Cheetahs and lions compete for food, territory, and other resources.
Saying they compete to “occupy” possible numerical population sizes is idiosyncratic at best.

117
species... will continue to live in a certain state of equilibrium. (Gause
1934, 98)
But nowhere did Gause explain how dierent niches must be to ensure coexis-
tence, how niches could be individuated, or, most importantly, how niche con-
siderations could help determine the competition coecients required by the
Lotka- Volterra competition equations. at dierent species typically utilize dif-
ferent food sources was well known well before Darwin, and existing models of
competition had already captured the resulting interspecic dynamics with mathe-
matical precision (e.g., Lotka 1932). Rather than constitute the indispensable con-
ceptual core of Gause’s work supporting CEP, his allusions to the niche seem more
gloss than grist.
3. HUTCHINSON’S N DIMENSIONAL HYPERVOLUME
For many ecologists, Hutchinson’s denition in “Concluding Remarks” (1957) was
a watershed moment:
it was not until Hutchinson’s “Concluding Remarks” that the niche con-
cept was rigorously dened and its relationship to competition and
species diversity rigorously explored... Hutchinson succeeded in com-
bining both the Eltonian and Grinnellian concepts of niche into one
model. (Real and Levin 1991, 180–81)
is “revolutionary” account (Chase and Liebold 2003; Schoener 1989) set the tra-
jectory of niche- based theorizing in ecology for several decades.
Hutchinson’s denition, which rst appeared in a footnote of an earlier lim-
nological paper (Hutchinson 1944),
10
characterizes two concepts, a species’ funda-
mental and realized niche. For a specic species S
1
, consider all the environmental
variables x
1
, x
2
. .. x
n
that aect S
1
, which Hutchinson (1957, 416) emphasized
includes both biological and (nonbiological) physical factors. If these variables are
conceptualized as axes, they dene an abstract n- dimensional space. e subset
of this space in which S
1
can persist indenitely (i.e., have positive tness) is the
fundamental niche of S
1
, with an important qualication: persistence is assessed
in the absence of all competing species. Not all other species are excluded in this
assessment, as is sometimes incorrectly claimed, because some of the environ-
mental variables that dene the space are in fact species (e.g., S
1
’s food resources,
or obligate symbionts). Of course, species oen do face competitors that constrict
their range. e subset of the fundamental niche actually realized by S
1
given com-
petitive dynamics is its realized niche.
10. “e term niche (in Gause’s sense, rather than Elton’s) is here defined as the sum of all the envi-
ronmental factors acting on the organism; the niche thus defined is a region of an n- dimensional
hyper- space” (Hutchinson 1944, 20, n. 5).

118
In signicant ways, Hutchinson’s account breaks sharply with earlier work.
Unlike Grinnell’s niche but similar to Elton’s, the fundamental and realized niches
are abstractions, not portions of real- world environments. Any Hutchinsonian
niche might correspond to highly disjoint sets of areas in the real world. e sole
gure in “Concluding Remarks” illustrates this relationship, ‘biotop space’ being
the actual environment of the two species (see Fig. 1).
Figure 3. Two fundamental niches dened by a pair of variables in a two-dimensional niche
space. Only one species is supposed to be able to persist in the intersection subset region.
e lines joining equivalent points in the niche space and biotop space indicate the
relationship of the two spaces. e distribution of the two species involved is shown on the
right hand panel with a temperature depth curve of the kind usual in a lake in summer.
Abstraction can make empirical concepts less tractable, and Elton’s concept
is oen contrasted unfavorably with Grinnell’s in this regard (Griesemer 1992).
But Hutchinson’s abstract denition is coupled with a signicant conceptual shi
that greatly enhances tractability: niches are strictly dened in relation to species,
persistence of the latter determining the boundaries of the former (for fundamen-
tal niches). By denitionally tethering niches to species, Hutchinson’s account
is much clearer about how niches are individuated: positive tness delimits the
niche- relevant portion of a species’ causal nexus. Concerns about how niches are
delineable that plagued Grinnell and Elton’s conceptions don’t gain nearly the same
purchase. It may be exceedingly empirically dicult to ascertain, but in principle
at least it is clear how species’ niches can be delimited.
11
11. Note that what allows determination of niche boundaries is the focus on a particular species
property aected by its environment, positive tness. at focus precisies but it also narrows
scope. If members of a species stray from their domain of nonnegative tness and have signicant
impacts on other species or the abiotic environment, the source of those causal impacts would
seem to fall outside the purview of Hutchinsonian niche considerations.

119
Gains in precision and tractability, however, came with signicant costs.
Hutchinson himself highlighted some shortcomings. e denition assumes all
points comprising the hypervolume entail equal probability of persistence (1957,
417). In reality, (absolute) tness will vary markedly in any plausibly realistic
assessment. Capturing these important dierences requires explicit representation
of the relevant ecological dynamics; i.e., the more ne- grained causal details that
determine whether and how species persist. e denition also assumes all the
environmental variables characterizing the space can be linearly ordered, which
Hutchinson admitted, “In the present state of knowledge this is obviously not
possible” (1957, 417). It is not entirely clear what precisely the diculty is, and
Hutchinson did not elaborate. Schoener (1989, 90) mentioned prey species and
vegetation type as examples of not linearly orderable environmental variables,
without further explanation. It seems, however, that prey species can be ordered
by their abundance, or frequency of encounter. If particular vegetation types are
required for species persistence, and their existence is binary and not a matter of
degree, then these environmental variables would not be linearly orderable. But
such bivalence seems quite unrealistic. As habitats, patches of vegetation of dier-
ent types presumably come in dierent degrees of suitability for dierent species.
Suitably then seems to impose an ordering, from the optimally tness enhancing
to the barely positive- tness maintaining. But again, this task faces the necessity of
representing ne- grained causal details discussed immediately above.
Hutchinson also stipulated, but apparently did not perceive as problematic,
that the environmental variables were independent and thus determined spaces
with orthogonal axes. But this assumption is false in most cases. In the limnologi-
cal systems for which Hutchinson rst formulated the denition, for example, tem-
perature, nutrient availability, light penetration, and other variables impacting
species are all dependent on depth. Temperature and precipitation are interrelated
for most if not all ecological systems. Nonindependence does not prevent construc-
tion of an abstract niche space, but it necessitates nonorthogonal, skewed axes and
coordinate systems to do so. Besides making visualization much more dicult, it
also renders some techniques used to analyze the detailed dynamics upon which
niche spaces depend inapplicable (e.g., the Fourier method for partial dierential
equations representing those dynamics).
ese are nontrivial problems, but they pale in comparison with the limitation
imposed by the signicant conceptual shi away from Grinnell and Elton’s account:
dening niches in terms of species persistence. at relativization, for instance,
makes the notion of a vacant niche conceptually incoherent.
12
A niche absent an
occupying species is impossible because the former is denitionally dependent on
the latter. And without the notion of a vacant niche— and in general a substan-
tive, species- independent niche concept— Hutchinson’s account is explanatorily
12. Hutchinson sometimes failed to recognize this implication of his denition (e.g. 1957, 424; 1959,
150; 1978, 161).

120
impoverished. It does not have the resources to explain phenomena such as eco-
logical equivalents, adaptive radiation into novel environments, similarities of
dierent communities’ structure, and others that were squarely in Grinnell and
Elton’s purview. e idea that “the Grinnellian niche and the Eltonian niche are
united through correspondence between points in N [the abstract niche space]
and points in B [the biotop space]” (Real and Levin 1991, 181) in Hutchinson’s
denition just fails to recognize how dramatically his concept diverges from theirs,
and how that divergence impacts its explanatory capabilities.
For the same reason, the Hutchinsonian niche cannot ground or otherwise be
the basis of the competitive exclusion principle. As indicated above, competitive
dynamics are excluded from the fundamental niche’s characterization. e real-
ized niche, on the other hand, assumes the principle: “we should expect that, in the
part of the hyperspace where the overlap occurred, competitive exclusion would
take place and the overlap would either be incorporated into the niche of one or
the other species or be divided between the two, producing the realized or post-
interactive niches of the two species” (Hutchinson 1978, 159).
13
Hutchinson, along
with many ecologists at the time, thought competition was the primary driver of
community structure; he called the CEP “a principle of fundamental importance”
(1957, 417). Hutchinson’s niche concepts reect this commitment, they don’t inde-
pendently support it.
14
Niches as dened by Hutchinson therefore convey very little information about
community dynamics. Hutchinsonian niche considerations cannot, for example,
determine which of two competitors will outcompete the other, and to what degree.
In fact, answering most questions that ecologists nd important— that competi-
tive exclusion will occur at all when fundamental niches overlap (without simply
assuming it will), how it occurs, the dimension size n of the hyperspace, the identity
13. ere are technical complications in assessing the relevant “overlap” that expose further chal-
lenges of Hutchinson’s hypervolume approach. First, if such overlap could be assessed in niche
space, it would require determining the intersection of two subspaces in a more expansive space
dened by the total set of environmental variables for both species. e fundamental niches of
dierent species will almost always be dened with nonidentical sets of environmental variables,
hence the need for the more expansive superspace. Although dierent species are sometimes
similar in specic ways, which can generate competition, they almost always have signicantly
dierent ecological requirements and tolerances in other ways. But, second, and more impor-
tantly, it is unclear overlap can even be assessed in that abstract space. When competition occurs,
it occurs in the real- world space species physically occupy (the “biotop” space). Trees compete for
light and soil nutrients in geographically coincident portions of the rainforest; pelagic birds com-
pete for nesting sites in specic clis of remote ocean islands. Competitive exclusion in the biotop
space would then translate into exclusion in portions of the abstract (nonspatially explicit) niche
space (see Fig. 1) that do not intersect in any set- theoretic or geometric sense. Simply talking of
overlap in an abstract hyperspace is therefore not an adequate representation of (spatially explicit)
competitive dynamics.
14. at commitment, furthermore, reects but one view of what primarily governs the structure of
biological communities. Fundamental and realized niches could be dened, for instance, with
predation, mutualism, or other ecological interactions at the forefront. If competition is not the
main driver of patterns and processes in the ecological world, Hutchinson’s approach will miss
much of what does.

121
of those dimensions, etc.— requires an explicit representation of species dynamics.
As a tool for representing and thereby understanding the dynamics responsible for
community structure and species properties, the Hutchinsonian niche is hardly
the epoch- making innovation it is oen heralded as.
4. CONCLUSION
ese shortcomings of past accounts are not a historical curiosity. In a recent eort
to reinvigorate niche- based ecological theorizing aer Hubbell’s (2001) inuential
neutral theory, Chase and Leibold review past characterizations of the concept and
propose a denition aiming at synthesis:
NICHE DEFINITION #1: the joint description of the environmental
conditions that allow a species to satisfy its minimum requirements so
that the birth rate of a local population is equal to or greater than its
death rate along with the set of per capita eects of that species on these
environmental conditions. (2003, 15)
e next sentence claries the denition is “a simple joining of the two concepts
that we have outlined in our historical review,” by which they mean a Grinnellian-
Hutchinsonian concept and Eltonian one.
15
In eect, then, this is a bipartite notion:
NICHE =
df
(i) all factors causally relevant to a species’ persistence;
(ii) all the species’ causal impacts on those factors.
But this characterization’s sweeping generality raises serious concerns about its
scientic utility. Rather than yielding something fruitful— that would, say, provide
guidance in representing and analyzing community dynamics— this niche de-
nition simply seems to acknowledge such dynamics exist. It’s as if, in a chemistry
context, one were told that the matter concept— with zero information about what
specic forms it can take, its compositional building blocks, or its connections
with other properties or lawful regularities— is nevertheless the key to chemical
analysis. What this niche denition oers seems largely to be a superuous gloss
on the causal details actually required to assess species persistence.
Chase and Leibold don’t explicitly recognize this deciency, of course, but
they may suspect something is amiss about the rst denition because a page later
they oer a second they claim is more precise:
NICHE DEFINITION #2: the joint description of the zero net growth
isocline (ZNGI) of an organism along with the impact vectors on that
15. at’s inaccurate. Any partitioning of environments independent of species persistence consid-
erations, Grinnell’s conception, is absent. Species’ functional properties that cannot be charac-
terized relative to environments in any straightforward way, Elton’s conception, are also absent.
Rather, this seems largely to be a recasting of Hutchinson’s notion.

122
ZNGI in the multivariate space defined by the set of environmental
factors that are present. (2003, 16)
‘Zero net growth isocline’ is short for the population values where
Revealingly, just before DEFINITION 2 Chase and Leibold (2003) say, “we will
use simple population dynamics models to justify a second more precise version
of this denition [#1].” In other words, niches are only determinable, and this de-
nition is only justiable, once species interactions have already been represented
in those models. at is, this niche concept itself contributes little or nothing to
determining that representation.
In the rest of their book this prioritization is never upended. It contains inter-
esting extensions of resource utilization models rst developed by Tilman (1982)
and sophisticated analyses of how empirical data might bear on them. But one
despairs of nding any nonredundant contribution the niche concept, as they dene
it, makes to these analyses. e absence isn’t surprising. Tilman’s (1982) highly inu-
ential book mentions ‘niche’ exactly four times, and in each case it refers to a label
used by others.
e shortcomings described above don’t impugn the sophisticated modeling
and empirical studies done under a ‘niche’ rubric. But they do indicate there isn’t
some insightful and foundational concept at the base of this work, in some way
guiding and shaping it all. at work stands alone.
REFERENCES
Chase, J., and M. Leibold. 2003. Ecological Niches. Chicago: University of Chicago Press.
Crowcro, P. 1991. Elton’s Ecologists. Chicago: University of Chicago Press.
Elton, C. 1927. Animal Ecology. Chicago: University of Chicago Press.
———. 1954. “An Ecological Textbook.” Journal of Animal Ecology 23: 282–84.
Gause, G. 1934. e Struggle for Existence. Baltimore, MD: Waverly Press.
Griesemer, J. 1990. “Material Models in Biology.” PSA Proceedings 2: 79–93.
———. 1992. “Niche: Historical Perspectives.” In Key Words in Evolutionary Biology, edited by E. Keller
and E. Lloyd, pp. 231–40. Cambridge, MA: Harvard University Press.
Grinnell, J. 1904. “e Origin and Distribution of the Chestnut- Backed Chickadee.” Auk 21:375–77.
———. 1917. “e Niche- Relationships of the California rasher.” e Auk 34: 427–33.
———. 1924. “Geography and Evolution.” Ecology 5: 225–29.
Haldane, J. B. S. 1924. “A Mathematical eory of Natural and Articial Selection: Part I.” Transactions
of the Cambridge Philosophical Society 23: 19–41.
Hardin, G. 1960. “e Competitive Exclusion Principle.” Science 131: 1292–97.
Hubbell, S. 2001. e Unified Neutral eory of Biodiversity and Biogeography. Princeton, NJ: Princeton
University Press.
Hutchinson, G. E. 1944. “Limnological Studies in Connecticut. Part 7. A Critical Examination of
the Supposed Relationship between Phytoplankton Periodicity and Chemical Changes in Lake
Waters.” Ecology 25: 3–26.
———. 1957. “Concluding Remarks.” Cold Spring Harbor Symposia on Quantitative Biology 22: 415–27.
———. 1959. “Homage to Santa Rosalia or Why Are ere So Many Kinds of Animals?” American
Naturalist 93: 145–59.
———. 1978. An Introduction to Population Ecology. New Haven, CT: Yale University Press.
123
Kingsland, S. 1995. Modeling Nature. 2nd edition. Chicago: University of Chicago Press.
Lawton, J. 1982. “Vacant Niches and Unsaturated Communities: A Comparison of Bracken Herbivores
at Sites on Two Continents.” Journal of Animal Ecology 51: 573–95.
Lotka, A. 1932. “e Growth of Mixed Populations: Two Species Competing for a Common Food
Supply.” Journal of the Washington Academy of Sciences 22: 461–69.
Odenbaugh, J. Forthcoming. “Neutrality, Niche, and Nulls: Causal Relevance in Ecology.” In Philosophical
Perspectives on Causal Reasoning in Biology, edited by C. Waters and J. Woodward. Minneapolis:
University of Minnesota Press.
Pearce, T. 2010. “ ‘A Great Complication of Circumstances’— Darwin and the Economy of Nature.”
Journal of the History of Biology 43: 493–528.
Peters, R. 1991. A Critique for Ecology. Cambridge: Cambridge University Press.
Real, L., and S. Levin. 1991. “e Role of eory in the Rise of Modern Ecology.” In Foundations of
Ecology, edited by L. Real and J. Brown, pp. 177–91. Chicago: University of Chicago Press.
Schoener, T. 1989. “e Ecological Niche.” In Ecological Concepts: e Contribution of Ecology to an
Understanding of the Natural World, edited by J. Cherrett, pp. 79–113. Oxford: Blackwell.
Tilman, D. 1982. Resource Competition and Community Structure. Princeton, NJ: Princeton University
Press.
