

OTTO KINNE
Editor

The late Frank H. Rigler
and Robert H. Peters
SCIENCE AND LIMNOLOGY
Introduction (Otto Kinne)
Frank H. Rigler and Robert H. Peters: A Laudatio
(Jürgen Overbeck)
Publisher: Ecology Institute
Nordbünte 23, D-21385 Oldendorf/Luhe
Germany
E
C
O
L
O
G
Y
I
N
S
T
I
T
U
T
E

Robert H. Peters
Department of Biology
McGill University
Montreal, PQ
Canada H3A 1B1
ISSN 0932-2205
Copyright © 1995, by Ecology Institute, D-21385 Oldendorf/Luhe, Germany
All rights reserved
No part of this book may be reproduced by any means, or transmitted, or translated without
written permission of the publisher
Printed in Germany
Typesetting by Ecology Institute, Oldendorf
Printing and bookbinding by Konrad Triltsch, Graphischer Betrieb, Würzburg
Printed on acid-free paper

To Ann and Antonella


Contents
Introduction (Otto Kinne) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . XI
Frank H. Rigler and Robert H. Peters: A Laudatio (Jürgen Overbeck) . . . . . . XXI
Prefaces . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . XXIII
Prologue . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
I WHY READ ABOUT SCIENCE? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
Some Misrepresentations of Science . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
Some Basic Distinctions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
Facts and theories . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
Induction and deduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
Empirical and explanatory theories . . . . . . . . . . . . . . . . . . . . . . . . . 15
The Importance of Science . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16
The Growth of Science . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
Summary . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
II A BRIEF HISTORY OF METHOD . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
Three Ways to Knowledge . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
Aristotle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22
Deduction and Induction in the Age of Reason . . . . . . . . . . . . . . . . . . . 25
Logical Positivism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28
On causality . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28
On new ideas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30
Sir Karl Popper . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
III NORMAL SCIENCE AND PSEUDO-SCIENCE . . . . . . . . . . . . . . . . . . 35
Kuhn’s “Normal” Science . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35
An historical model of science . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36
“Pseudo-Science” . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
Velikovsky . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
IV THE ECOLOGISTS’ DISEASE: TWO PERSONAL EXAMPLES . . . . . 47
Science and Ecology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
Non-theories . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49
The niche . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49
The competitive exclusion principle . . . . . . . . . . . . . . . . . . . . . . . . 50
Weak Theories . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 54
Evolution by natural selection . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 54
Concepts and measurement of phosphorus fractions . . . . . . . . . . . . 56
V BROADER SYMPTOMS OF THE ECOLOGISTS’ DISEASE . . . . . . . . 63
Framing Scientific Proposals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
The Reception of Moderately Restrictive Theories . . . . . . . . . . . . . . . . 64

The Pursuit of Ecological Concepts . . . . . . . . . . . . . . . . . . . . . . . . . . . 67
The limiting factor . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 68
Unconcern . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 70
Multiple limitation in the sea . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 70
Inattention to Detail . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 72
The calculation of secondary productivity . . . . . . . . . . . . . . . . . . . . 73
Some Consequences . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 77
VI WHY LIMNOLOGY? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 79
What is Limnology? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 79
What is Science? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 80
Ecological Theories . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 81
The ecosystem concept . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 82
Why limnology? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83
A paradigm shift in limnology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87
Limnology and marine science . . . . . . . . . . . . . . . . . . . . . . . . . . . . 91
Why limnology — an answer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 93
VII REDUCTIONISM VERSUS HOLISM:
AN OLD PROBLEM REJUVENATED BY THE COMPUTER . . . . . . . 95
The Place of Philosophical Debates in Biology . . . . . . . . . . . . . . . . . . . 95
Malloy and the principle of trim . . . . . . . . . . . . . . . . . . . . . . . . . . . 96
Vitalism and mechanism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 97
Organicism and holism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 98
Holism and reductionism in ecology . . . . . . . . . . . . . . . . . . . . . . . . 100
What is Systems Analysis? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 101
Some problems with proposed solutions . . . . . . . . . . . . . . . . . . . . . 104
Two Personal Experiences . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 106
The Char Lake Project . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 106
Zooplankton feeding . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 110
The Reality of Systems Analysis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 113
Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 114
VIII SOURCES OF ECOLOGICAL CREATIVITY . . . . . . . . . . . . . . . . . . . . 117
The Challenge of Creativity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 117
The Existing Literature as an Inspirational Device . . . . . . . . . . . . . . . . 119
Dissection . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 120
Mechanism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 121
Dichotomies and categories . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 121
Analysis of variance . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 122
Extensions, additions and modifications . . . . . . . . . . . . . . . . . . . . . 122
Technologies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 123
Complications . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 123
The Danger of Conventionalism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 125
Creative Alternatives for Normal Ecology . . . . . . . . . . . . . . . . . . . . . . 126
Syllogisms and analogies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 126
A return to application . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 127
CONTENTS

IX EMPIRICAL LIMNOLOGY . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 129
Social Demands and Scientific Supply . . . . . . . . . . . . . . . . . . . . . . . . . 129
Pessimists and Optimists . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 130
Testing the alternatives . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 131
Holists and reductionists . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 132
What to Predict? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 133
A Research Program in Holistic Empirical Ecology . . . . . . . . . . . . . . . 134
How green is my lake? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 134
Phosphorus concentration . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 136
The growing school of empirical limnology . . . . . . . . . . . . . . . . . . 143
Summary — A Future for Ecology . . . . . . . . . . . . . . . . . . . . . . . . . . . . 147
X AN EDUCATION IN SCIENCE: EVALUATION . . . . . . . . . . . . . . . . . . 149
On Advising Teachers . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 149
The Goals of a University Education in Science . . . . . . . . . . . . . . . . . . 150
Strategies for Teaching . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 151
Empowerment by theory . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 152
Understanding through explanation . . . . . . . . . . . . . . . . . . . . . . . . . 153
Paradigmatic indoctrination . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 155
Disciplinary description . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 155
An Evaluation of Teaching in Biology and Ecology . . . . . . . . . . . . . . . 156
Nurture or nature . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 156
The problem with textbooks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 157
The problem with courses . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 159
Repercussions for graduate training . . . . . . . . . . . . . . . . . . . . . . . . . 160
A lesson from the literature . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 161
XI AN EDUCATION IN SCIENCE: PRESCRIPTIONS . . . . . . . . . . . . . . . . 163
The Undergraduate Program . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 163
The problem of confidence . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 164
De-enrichment and dis-integration . . . . . . . . . . . . . . . . . . . . . . . . . . 166
Hierarchical themes for undergraduate education . . . . . . . . . . . . . . 168
A theoretical typology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 170
Graduate Education . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 171
The importance of role models . . . . . . . . . . . . . . . . . . . . . . . . . . . . 171
Wise choices in graduate education . . . . . . . . . . . . . . . . . . . . . . . . . 172
Administrative Advice . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 175
XII THE QUESTIONS OF RELEVANCE . . . . . . . . . . . . . . . . . . . . . . . . . . . 177
What Use is Science to Society? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 177
Does science differ from applied research and technology? . . . . . . 178
Does Science Merit Support? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 180
How can we evaluate our science? . . . . . . . . . . . . . . . . . . . . . . . . . . 180
How can ecology merit support? . . . . . . . . . . . . . . . . . . . . . . . . . . . 185
CONTENTS

XIII FUNDING DECISIONS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 187
The Central Problem for Research Funding . . . . . . . . . . . . . . . . . . . . . 187
Reasonable expectations from research . . . . . . . . . . . . . . . . . . . . . 188
How to gamble with research funds . . . . . . . . . . . . . . . . . . . . . . . . 191
Some realities of ecological research . . . . . . . . . . . . . . . . . . . . . . . 194
The Dream of Multi-Disciplinary Environmental Science . . . . . . . . . . 195
The advantages of team research . . . . . . . . . . . . . . . . . . . . . . . . . . 195
The problems of multi-disciplinary research in ecology . . . . . . . . . 196
Where do we go from here? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 200
XIV DARWIN AND EVOLUTIONARY SCIENCE . . . . . . . . . . . . . . . . . . . . 201
Darwin on the Galapagos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 202
Critics of Darwin . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 204
The first school: early emotionals . . . . . . . . . . . . . . . . . . . . . . . . . . 204
The second school: directional deists . . . . . . . . . . . . . . . . . . . . . . . 207
The third school: cataclysmic creationists . . . . . . . . . . . . . . . . . . . 208
The fourth school: Popperian purists . . . . . . . . . . . . . . . . . . . . . . . 209
Two other biological schools . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 211
Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 211
XV IS THE FUTURE GRIM? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 213
The Gilt Age of University Research . . . . . . . . . . . . . . . . . . . . . . . . . . 213
The Gathering Challenge to University Science . . . . . . . . . . . . . . . . . . 214
University Responses . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 216
A Policy for the Future: Closing the Aspiration Gap . . . . . . . . . . . . . . 217
Undergraduate teaching . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 217
Graduate training . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 219
Scholarship . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 219
Administration . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 220
Two reservations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 221
Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 222
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 223
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 225
CONTENTS

Introduction
Otto Kinne
Ecology Institute, Nordbünte 23, D-21385 Oldendorf/Luhe, Germany
Science and Limnology is likely to become a milestone in ecological reason-
ing and research. Based to a considerable extent on unpublished thoughts
and notes of Frank H. Rigler, and written by the winner of the Ecology Insti-
tute Prize 1991 in the field of limnetic ecology, Robert H. Peters of McGill
University, Montreal, Canada, the book is enlightening, challenging and
provocative. It is enlightening, because it views science in general and ecol-
ogy in particular from unconventional angles; it is challenging, because it
criticizes many of the ways in which ecologists think and approach their sub-
jects; it is provocative, because the author presents an unattractive picture of
present-day ecology and a harsh assessment of university education and
research. Above all, Science and Limnology is a worthy companion to
previous Excellence in Ecology books: it offers the well-written, concise and
easy-to-read personal views of an outstanding performer in his field of
expertise. Rob Peters has written this book as a professor in the best sense of
that word. He professes his insights, beliefs and convictions with courage
and honesty, and he considers his topics with care and a keen mind. His
views reach far beyond the horizon suggested by the book’s title, far into the
realms of science history, philosophy and methodology; into the relevance of
science to society; and into the centers where science is at home and where
scientists are formed — the universities.
The essence of the message conveyed by Peters is this: ecologists have
collected impressive amounts of observations and facts, but they have failed
to sufficiently identify and formulate theories that go beyond the facts —
theories that can be tested and that can predict. To aid in solving the many
problems which press on modern human societies and in controlling and
restricting the ever-increasing deformation of nature, ecological research
must focus, more so than in the past, on empirical, holistic approaches that
facilitate prediction. Peters insists that the failure of ecologists to produce
useful predictions is not a consequence of the complexity of their subjects,
but of the complexity of their approaches. He believes that his call for a more
empirical, holistic strategy will be heard and accepted by ecologists and that
it will help mankind to preserve itself and to save its environment.

XII INTRODUCTION

For Peters the essence of science is the creation, testing and use of theory.
In the process of creating theory, induction plays an important role. It is here
that inspiration and intuition enter the scene. While theory is the backbone of
research, it can never be beyond doubt. Theories must be tested time and
again. The prediction must be compared to further observation. Since scien-
tific knowledge always remains hypothetical, the ultimate arbiter of scien-
tific research is observation. ‘The main goals of science are to make theo-
ries, to use theories to make predictions and to assess those predictions
against observation’ (p. 21).
Peters defines ecology ‘as the science that predicts the abundance, distri-
bution and other characteristics of organisms in nature’ (p. 81). He deplores
that ecologists have failed to appreciate the nature of their science: ‘... much
of ecology is confused in its goals, uncertain of its thoughts, and inconsistent
in its terminology.’ Chapter V portrays ecologists ‘as nonchalant about their
tests, careless in their measurements, yet closed-minded in considering alter-
natives’ (p. 77).
Turning to his main subject, limnology, Peters points out that the leading
role this branch of science once played in ecological research was lost,
largely because the assumption that ecological theory required an isolated
microcosm was discovered to be a misapprehension. Based on the tools and
information produced over many decades, limnologists are now turning to
empirical theories that predict. Thus, modern limnology is becoming a
leader again, this time showing the way towards predictive ecology.
Examining university research and teaching, Peters identifies shortcom-
ings at every level, but he also makes suggestions for improvements and
offers some practical advice. In regard to teaching science, he stresses again
our failure to appreciate the significance and nature of theory. Reconsidering
and contemplating science should lead to profound changes in research and
in the ways professors teach students: ‘Too much research is done for the
same reason that a mountain is climbed (“because it is there”), and too little
time is spent questioning the motives for doing so’ (p. 179). On the other
hand, deplores Peters, big science and societal power have separated profes-
sors from their ideals and goals, and thus almost destroyed the university as
an intellectual retreat.
XIIIINTRODUCTION
Ecology Institute Prize 1991 in the field of limnetic ecology. Reproduction of the prize
awarding document

Excellence in Ecology Books
Published bythe International EcologyInstitute (ECI), thebook series “Excel-
lence in Ecology” (EE) is made available at cost price.* EE books are also
donated to scientific libraries in Third-World countries. The books are
authored by recipients of the Ecology Institute Prize. This prize honors the
sustained high performance of outstanding research ecologists. Prizes are
awarded annually, in a rotating pattern, for the fields of marine, terrestrial and
limnetic ecology. Laureates are selected by a jury of seven ECI members
appointed by the director. EE books offer the laureates the chance to publish
theirpersonal views onthestate oftheart intheirfieldsof expertiseand tobring
to the attention of a world-wide audience their insights into the knowledge,
problems and realities that form the biological basis for human existence.
EE books address fellow scientists, teachers, students and decision
makers who must translate ecological information into practicable rules and
laws for the benefit of nature and mankind.
The aims and activities of the Ecology Institute have been outlined in
EE Book 3 (pp. VIII–IX).
In addition to the ECI Prize, the ECI awards an annual IRPE Prize (Inter-
national Recognition of Professional Excellence) which honors a young (not
more than 40 years of age) research ecologist who has published uniquely
independent and/or challenging papers representing an important scientific
breakthrough and/or who must work under particularly difficult conditions.
The ECI also supports, via the Otto Kinne Foundation (OKF), promising
postgraduates in environmental sciences in East European countries — espe-
cially in the fields of ecology, diseases of organisms, and climate research.
The OKF aids postgraduates — without distinction of race, religion, nation-
ality or sex — by providing financial assistance for professional travel, sci-
entific equipment or published information. For details write to the President
of the Foundation: Dr. J. Lom, Institute of Parasitology, Academy of
Sciences of the Czech Republic, Branisˇovská 31, 370 05 C
ˇ
eské Budeˇjovice,
Czech Republic; tel. (+42) 38 41158, fax (+42) 38 47743.
XIV INTRODUCTION
*Address orders for EE books to:
*Ecology Institute, Nordbünte 23, D-21385 Oldendorf/Luhe, Germany
Payment may be made via credit card (American Express, Visa, Euro/ Mastercard;
please give account number and expiration date).
An order for the whole series is accepted at a 10% reduced price.
For book authors, titles and prices, consult pp. XV–XVII.

Nominations for ECI and IRPE Prizes (accompanied by CV, list of publi-
cations, and a statement why, in the opinion of the nominator, the nominee
qualifies for the prize) are invited from research ecologists on a global scale.
They should be sent to the chairperson of the respective ECI Jury, or, alter-
natively, to the ECI’s director, who will then forward them to the chair-
person. Eligible are all ecologists engaged in scientific research (except the
ECI’s director, the Jury’s chairperson, and previous Laureates; Jury mem-
bers nominated will be replaced by other ECI members). The Jury selects
prize winners using the nominations received as well as their own knowledge
of top performers and their own professional judgement.
Nominations for OKF Fellows, addressed to Dr. J. Lom (see above) and
accompanied by a letter of support as well as a documentation of the
nominees’ performance, are invited from ECI members and members of the
Editorial Staffs of the three international Inter-Research journals, Marine
Ecology Progress Series, Diseases of Aquatic Organisms, and Climate
Research.
ECI Prize Winners, Their Major Scientific Achievements
and Their Books
Tom Fenchel (Helsingør, Denmark), ECI Prize winner 1986 in marine ecology.
Quotation of the Jury (Chairman: John Gray, Oslo, Norway)
The Jury found Professor T. Fenchel’s contribution to ecological knowledge in a variety of
research fields to be of the highest international class. In particular, the Jury cites his brilliant
and uniquely important studies on the microbial loop which have opened up a fundamentally
new research field. Professor Fenchel is, in addition, an excellent publicizer in his field of
research with authorship of a number of standard works in marine ecology.
Book 1: Ecolo
gy – Potentials and Limitations. (Published 1987; price DM 67 plus postage
and handling)
Edward O. Wilson (Cambridge, MA, USA), ECI Prize winner 1987 in terrestrial ecology.
Quotation of the Jury (Chairman: Sir Richard Southwood, Oxford, UK)
Professor E. O. Wilson is distinguished for his many contributions to different aspects of ecol-
ogy and evolutionary biology. His life-time love of Nature, a theme explored in his book “Bio-
philia”, has been particularized in his study of ants leading to major new insights on the evo-
lution of castes and the operation of social systems. His seminal “Sociobiology”, derived
from this work, has founded a new branch of science, between ecology and the social sci-
ences. With the late Robert MacArthur he was the originator of the modern theories of island
biogeography that have contributed not only to the understanding of island biota, but to com-
munity and population ecology.
Book 2: Success and Dominance in Ecosystems: T
he Case of the Social Insects. (Published
1990; price DM 49 plus postage and handling)
XVECI PRIZE WINNERS

Gene E. Likens (Millbrook, NY, USA), ECI Prize winner 1988 in limnetic ecology.
Quotation of the Jury (Chairman: William D. Williams, Adelaide, Australia)
Gene Likens is a distinguished limnologist who has made salient contributions to many fields
of limnology. In 1962 he initiated and developed (with F. H. Bormann) the Hubbard Brook
Ecosystem Study in New Hampshire. Comprehensive investigations in this study provided a
model for ecological and biogeochemical studies worldwide. A major finding of the study was
that rain and snow are highly acidic. “Acid rain” is now recognized as one of the major envi-
ronmental hazards in North America, Europe and elsewhere. Elected to the American
Academy of Sciences in 1979, and the National Academy of Sciences in 1981, Gene Likens is
a highly worthy recipient of the 1988 ECI Prize in Limnetic Ecology.
Book 3: T
he Ecosystem Approach: Its Use and Abuse. (Published 1992; price DM 59 plus
postage and handling)
Robert T. Paine (Seattle, WA, USA), ECI Prize winner 1989 in marine ecology.
Quotation of the Jury (Chairman: Tom Fenchel, Helsingør, Denmark)
Robert T. Paine has made substantial and original contributions to marine biology and to
ecology in general. In particular the Jury mentions the discovery of the role of patch forma-
tion and properties of food web structure in shaping communities of sedentary organisms.
These studies (of which several have become classics of marine ecology) have fundamentally
changed the way in which we view marine benthic communities. This work has also served as
an inspiration for innovation in the mathematical description of community processes and has
had a lasting impact on our understanding of “landscape dynamics”, of equal importance to
the development of the science of ecology and to conservation ecology.
Book 4: Mar
ine Rocky Shores and Community Ecology: An Experimentalist’s Perspective.
(Published 1994; price DM 59 plus postage and handling)
Harold A. Mooney (Stanford, CA, USA), ECI Prize winner 1990 in terrestrial ecology.
Quotation of the Jury (Chairman: John L. Harper, Penmaenmawr, UK)
Professor Harold A. Mooney is distinguished for his studies of the physiological ecology of
plants, especially of arctic-alpine and mediterranean species. He has explored the ways in
which plants allocate carbon resources and expressed this allocation in terms of costs, bene-
fits and trade-offs. This has given a quantitative dimension to the study of plant-animal inter-
actions and acted to integrate physiological ecology with population biology, community
ecology, and ecosystem studies.
Book 5: T
he Globalization of Ecological Thought. (To be published soon)
Robert H. Peters (Montreal, PQ, Canada), ECI Prize winner 1991 in limnetic ecology.
Quotation of the Jury (Chairman: Jürgen Overbeck, Plön, Germany)
Professor R. H. Peters’ contributions to the fields of limnology and ecology have been numer-
ous and far reaching. His work on phosphorus cycling in lakes provides examples of excellent
research illuminating a number of important aspects regarding the movement and availabil-
ity of phosphorus in aquatic systems. His book “The Ecological Implications of Body Size”
gives a powerful overview of the utility of allometric relationships for the study of ecological
problems and for building ecological theory.
Book 6: Science and Limnolo
gy. (Published 1995; price DM 74 plus postage and handling.)
Authors: The late F. H. Rigler and R. H. Peters
XVI INTRODUCTION

Dr. David H. Cushing (Lowestoft, United Kingdom), ECI Prize winner 1992 in marine
ecology.
Quotation of the Jury (Chairman: John Costlow, Beaufort, NC, USA)
Dr. David H. Cushing has, for many years, made an enormous contribution to the field of
marine ecology through his numerous publications and his original ideas. His work continues
to be highly influential in fisheries and plankton ecology. Although first published over ten
years ago, his pioneering studies on the dynamics of a plankton patch, the feeding of cope-
pods, the ‘match-mismatch’ theory of recruitment and the climatic influences on plankton and
fisheries remain of central importance.
Book 7: Recr
uitment in Marine Fish Populations. (To be published 1995/96)
Paul R. Ehrlich (Stanford, CA, USA), ECI Prize winner 1993 in terrestrial ecology.
Quotation of the Jury (Chairman: Harold A. Mooney, Stanford, CA, USA)
Dr. Paul Ehrlich’s scientific contributions have been substantial and sustained. The quality
and depth of his interpretation of environmental issues to students, the general public, and to
policy makers is unrivaled. His concern for both environmental quality and environmental
justice has rarely been matched. He has made fundamental contributions to the study of pop-
ulation biology utilizing butterflies as a model system. These studies have had a large impact
on how we view the population structure of organisms and have provided important guide-
lines on the conservation of wild populations.
Book 8: A W
orld of Wounds: Ecology and Human Predicament. (To be published 1995/96)
IRPE Prize Winners and Their Major Scientific Achievements
Colleen Cavanaugh (The Biological Laboratories, Harvard University, Cambridge, MA
02138, USA), IRPE Prize winner 1986 in marine ecology.
Quotation of the Jury (Chairman: John Gray, Oslo, Norway)
The Jury found the research of Dr. C. Cavanaugh on chemosynthesis – initially concerning
hot-vent fauna but extended to other sulphide-rich habitats – to be highly original and to rep-
resent a major scientific breakthrough. Her hypothesis, formulated whilst a beginning gradu-
ate student, met severe opposition from established scientists with opposing views, but never-
theless proved to be correct. The Jury acknowledge Dr. Cavanaugh’s brilliant and
independent research in understanding chemosynthetic energetic pathways.
Karel
ˇ
Simek (Hydrobiological Institute, Czechoslovak Academy of Sciences, 370 05
ˇ
Ceské
Budˇejovice, Czechoslovakia), IRPE Prize winner 1991 in limnetic ecology.
Quotation of the Jury (Chairman: Jürgen Overbeck, Plön, Germany)
Dr. Karel Simek belongs to the generation of young limnologists in Eastern Europe who –
despite lack of international information exchange – published, under difficult conditions,
excellent contributions to the field of Aquatic Microbiology. He enjoys a high international
reputation. Under the present, improved conditions Simek is likely to proceed even more
successfully to new professional horizons.
XVIIIRPE PRIZE WINNERS

Richard K. Grosberg (Department of Zoology, University of California, Davis, CA 95616,
USA), IRPE Prize winner 1992 in marine ecology.
Quotation of the Jury (Chairman: John Costlow, Beaufort, NC, USA)
Richard K. Grosberg has not only published extensively on fundamental issues relating to
marine ecology, but has also demonstrated his understanding of marine ecology through
superb teaching of invertebrate zoology to undergraduate and graduate students. He is
acknowledged as a leader in adapting molecular techniques for the study of marine larvae
and in developing information on extraordinarily detailed mapping studies of the genetic
structure of adult populations of marine organisms.
Nikolai V. Aladin (Zoological Institute, Russian Academy of Sciences, St. Petersburg
199034, Russia), IRPE Prize winner 1993 in terrestrial ecology.
Quotation of the Jury (Chairman: Harold A. Mooney, Stanford, CA, USA)
Dr. Nikolai V. Aladin is one of Russia’s most eminent young ecologists. He has researched
environments in the former Soviet Union, particularly in Kazakhstan where he and a small
team have focussed upon the area of the Aral Sea. Dr. Aladin’s studies were performed dur-
ing a period of change, both in the patterns of organismic assemblages and in the political
structure of his country. These studies were undertaken in his own time and at his own
expense. It is only over the past few years that his studies have been officially supported and
their value recognized.
Ecology Institute Staff 1995 (in brackets: year of appointment)*
Director and Founder: Professor O. Kinne, D-21385 Oldendorf/Luhe, Germany
Marine Ecology
Terrestrial Ecology
XVIII INTRODUCTION
Prof. F. Azam, La Jolla, CA, USA (1985)
Prof. H.-P. Bulnheim, Hamburg, Germany (1984)
Prof. S. W. Chisholm, Cambridge, MA, USA
(1993)
Dr. D. H. Cushing, Lowestoft, UK (1993)
Prof. T. Fenchel, Helsingør, Denmark (1985)
Dr. N. S. Fisher, Stony Brook, NY, USA (1985)
Prof. J. Gray, Oslo, Norway (1984)
Prof. B.-O. Jansson, Stockholm, Sweden (1989)
Prof. V. Kasyanov, Vladivostok, Russia (1993)
Prof. E. Naylor, Menai Bridge, UK (1984)
Prof. S. W. Nixon, Narragansett, RI, USA (1989)
Prof. W. Nultsch, Hamburg, Germany (1994)
Prof. R. T. Paine, Seattle, WA, USA (1990)
Dr. T. Platt, Dartmouth, NS, Canada (1984)
Acad. Prof. G. G. Polikarpov, Sevastopol,
Ukraine (1985)
Dr. T. S. S. Rao, Bambolim, India (1985)
Prof. V. Smetacek, Bremerhaven, Germany (1993)
Prof. B. L. Wu, Qingdao, China (1993)
Acad. Prof. A. Zhirmunsky, Vladivostok, Russia
(1988)
*Following their receipt of the ECI prize, laureates are invited to join the institute’s staff
Prof. T. N. Ananthakrishnan, Madras, India
(1984)
Prof. F. S. Chapin, III, Berkeley, CA, USA
(1986)
Prof. J. Ehleringer, Salt Lake City, UT, USA
(1986)
Dr. P. Ehrlich, Stanford, CA, USA (1994)
Prof. M. Gadgil, Bangalore, India (1985)

Limnetic Ecology
Technical Staff (all Oldendorf/Luhe, Germany)
XIXECOLOGY INSTITUTE STAFF
Prof. I. Hanski, Helsinki, Finland (1993)
Prof. J. L. Harper, Penmaenmawr, UK (1986)
Prof. E. Kuno, Kyoto, Japan (1986)
Prof. A. Macfadyen, Coleraine, UK (1985)
Prof. H. A. Mooney, Stanford, CA, USA
(1991)
Dr. M. Shachak, Sede Boker, Israel (1989)
Acad. Prof. V. E. Sokolov, Moscow, Russia
(1986)
Prof. Sir R. Southwood, Oxford, UK (1986)
Prof. S. Ulfstrand, Uppsala, Sweden (1986)
Prof. E. O. Wilson, Cambridge, MA, USA (1988)
Prof. N. V. Aladin, St. Petersburg, Russia (1994)
Prof. J. I. Furtado, Washington, DC, USA (1985)
Prof. S. D. Gerking, Tempe, AZ, USA (1986)
Dr. J. E. Hobbie, Woods Hole, MA, USA (1986)
Dr. E. Kamler, Lomianki, Poland (1993)
Prof. W. Lampert, Plön, Germany (1993)
Prof. G. E. Likens, Millbrook, NY, USA (1989)
Prof. K. Lillelund, Hamburg, Germany (1985)
Prof. R. Margalef, Barcelona, Spain (1986)
Prof. J. Overbeck, Plön, Germany (1984)
Prof. T. J. Pandian, Madurai, India (1985)
Dr. E. Pattée, Villeurbanne, France (1987)
Prof. R. H. Peters, Montreal, PQ, Canada (1992)
Prof. E. Pieczyñska, Warsaw, Poland (1993)
Prof. J. G. Tundisi, São Paulo, Brazil (1990)
Dr. D. Uhlmann, Dresden, Germany (1989)
Prof. W. Wieser, Innsbruck, Austria (1987)
Prof. W. D. Williams, Adelaide, Australia (1986)
J. Austin
G. Bendler
M. Bruns
V. Cleary
C. Fesefeldt
R. Friedrich
B. Fromm
S. Hanson
R. Hooper
J. Hunt
H. Kinne
J. Kunert
M. Masuhr
T. Masuhr
W. Neel
R. Stedjee
H. Witt


Frank H. Rigler and Robert H. Peters:
A Laudatio
Jürgen Overbeck
Max-Planck-Institut für Limnologie, D-24302 Plön, Germany
With the sudden death of Professor Frank Rigler in 1982, we, his friends and
colleagues, lost a distinguished scientist and a leading limnologist with a
broad field of interest and research.
Who was Frank Rigler? Born in London in 1928, he received his Ph.D. in
limnology from the University of Toronto in 1954. He was married and had
5 children. Frank Rigler was Chairman of Biology at McGill University in
Montreal, Quebec, Canada, from 1976 to 1981. His special interests were
predicting the effects of nutrient enrichment on production in temperate and
subarctic lakes. His laboratory was also long concerned with the status of
ecology as a science and the role of ecological knowledge.
My first personal acquaintance with Frank Rigler was at the XIXth
International Limnological Congress in Winnipeg, Canada, in 1974, where
he gave a Plenary Lecture with the title Nutrient Kinetics and the New
Typology. This lecture provoked an extraordinarily controversial discussion.
Rigler began by saying that he doubted that studies on the details of the
phosphorus cycle were really advancing our knowledge at all. There already
existed an embarassingly large accumulation of facts in limnology. But sci-
entific advance comes only when we think up a new theory that overcomes a
difficulty experienced by the old theory. A qualified scientific theory must
be — in the sense of Karl Popper — potentially falsifiable. Starting from
this, Rigler presented, from a holistic point of view, nutritional-production
limnology using the ‘black-box’ approach, which may give us predictive
ability but no real understanding. Models are, by and large, purely empirical
descriptions of correlations between state variables. However, they pose
questions and the work of reductionists may suggest theories to answer these
questions. It was indeed an extraordinary lecture, quite different from the
usual way of presenting ecological results and systems.
Due to his early death, Rigler was unable to publish many of his ideas. In
this connection I will cite a letter of Professor M. L. Ostrofsky, Meadville,

Pennsylvania: “Rigler’s ideas about the nature of science, the requirements
of ‘good’ science, and the role of ecologists in shaping public issues are not
widely known. Rigler’s modesty prevented him from publishing many of his
ideas until late in his career. Many of Rigler’s ideas have been placed before
a larger scientific audience through the work of Dr. Robert Peters, Rigler’s
most articulate student, friend and colleague. It would be of enormous sig-
nificance if the full range of Rigler’s thoughts could be made accessible to a
larger audience. I cannot think of a more timely subject for a book; the recent
literature suggests that ecology is in the midst of a crisis of confidence and
identity. I cannot think of a more appropriate individual to undertake the
task.” The award of the 1991 ECI Prize in Limnetic Ecology to Robert Peters
now offers the unique opportunity for realizing this project. This book,
Science and Limnology, thus has two authors and two prefaces — a junior
author’s preface and a senior author’s preface. Robert Peters approached the
book as a “collaborative exercise, a collage in which Rigler’s enduring ideas
are set within a matrix of my own writing to produce a contemporary essay
about the science of limnology.” The basis of the 15 chapters is over 70 sets
of notes and lectures covering 25 years.
Robert Henry Peters was born in Toronto, Ontario, Canada, on August 2,
1946. He received his Ph.D. in 1972. His doctoral thesis on regeneration of
phosphorus by zooplankton — an issue which is still of great topical impor-
tance — was supervised by Frank Rigler. Highlights of Robert Peter’s
academic career include post-doctoral fellowships in Pallanza (Italy),
1972–1973; in Vienna (Austria), 1973; and in Munich (Germany), 1974. He
became Assistant Professor in 1974, Associate Professor in 1979 and Full
Professor in 1986 at McGill University. He has published 125 papers, notes
and reports, and authored a book on the ecological implications of body size.
His current research interests center on material flow in aquatic ecosystems,
zooplankton behavior, allometric relationships in autecology, and applied
environmental management.
I hope that Science and Limnology will be as inspiring, uncommon and
provoking for its readers as was the Plenary Lecture of Frank Rigler which I
had the pleasure to attend and discuss 20 years ago.
XXII LAUDATIO

Prefaces
Junior author’s preface
To win an international prize is a wonderful and surprising event. A hundred
other researchers could merit most prizes, and many worthy individuals go unre-
warded in any prize-giving. Merton (1968) has called these unrecognized but
worthy colleagues “occupants of the 41st chair”, in reference to the many tal-
ented individuals who never won the honour of a place in the Académie
Française, simply because the Académie is limited to 40 chairs. Deserving new-
comers must await the death of present occupants, and some die before an oppor-
tunity arises, as did Descartes, Molière, Pascal, and Diderot. Because I know the
unlikelihood of winning honours, the decision of the Ecology Institute to award
me its 1991 prize in limnetic ecology was first a surprise, and then a source of
great pleasure. It remains a reason for deep satisfaction and pride, as well as grat-
itude towards Professor Otto Kinne, Director of the Ecology Institute, Professor
Jürgen Overbeck, Chairman of the ECI Jury, the members of the ECI Jury, the
staff of the Institute, and the generous colleagues who lent me their support. Like
most critical scientists, I am prone to self-doubt, and therefore I am all the more
touched that not all of my ideas have been dismissed as either appallingly bad or,
worse, frightfully dull. I am also humbled to find myself in the ranks of previous
laureates whose contributions have been so much greater than my own.
I have an additional personal reason to be pleased with this prize. It has
allowed me to complete a project that I have wanted to do for over a decade: to
prepare the unpublished notes of Frank Rigler for publication. When Frank Rigler
died in 1982, the world of limnology lost one of its leading thinkers. The loss was
all the more tragic because most of his broader views on limnology, ecology and
science were never published. His influence had been a personal one, spreading
through conversation and occasional public lectures. As a summer employer and
undergraduate teacher, as my doctoral supervisor, as my long-time colleague and
close friend, and still as my conscience and model, Frank has been the major
influence on my career and scientific development. He has played a similar role
for other students, at levels ranging from undergraduate to senior scientist. For us
all, conversation with Frank was a treasure, a model of clear thought, logic, pen-
etration, relevance and simplicity. He taught us the nature of science, the power it
gives its followers, and the burdens of responsibility it places upon them. He was
a source of strength and inspiration, and he is sorely missed.
Rigler’s natural modesty stopped him from writing about his larger views for
most of his career. Soon after he finally began to write to a larger audience, his

voice was stilled by cancer. His few philosophical papers (Rigler 1975a, b,
1982a, b) were powerful and controversial, but they exposed only a fraction of his
ideas. Only those who knew his teaching at first hand, whether in university
courses, in scientific lectures, in panel discussions, or in quiet talks, realize how
much went unrecorded.
Rigler’s vision and virtues are even more needed now, but my aim in prepar-
ing this book is not simply to reproduce the text of his lectures and unpublished
notes. His notes were extensive and he was meticulous in preserving that mate-
rial, but even good lecture notes are not publishable as they stand. I have instead
approached the book as a collaborative exercise, a collage in which Rigler’s
enduring ideas are set within a matrix of my own writing to produce a contempo-
rary essay about the science of limnology. This cannot be the book Rigler would
have written, but because his spirit is so much a part of my everyday experience,
it may approach a book we could have written together, had he lived. For me, the
exercise has been profoundly rewarding. I have been able to reexamine the roots
of my own views and to appreciate the rich earth from which they developed. I
also discovered that many ideas I thought my own were actually foreshadowed in
his writings.
I have chosen to write in the first person singular, and therefore I have created
a non-existent author who is neither me nor Rigler, but some amalgam of us both.
Use of the first person preserves that conversational tone of Rigler’s lectures and
confirms this book as a personal document. Use of this chimeric “I” sometimes
results in incongruities since the pronoun clearly refers to only one of the two
authors, but on the whole I like the device. At times, it even allowed me to see my
ideas from a greater distance than I usually can.
The 15 chapters are based on over 70 sets of notes and lectures covering a
quarter century. I have recast and resorted most of the material, but most chapters
can still be appreciated separately. The book is arranged in a roughly chronolog-
ical order based on an intellectual ontogeny or autobiography whose main lines
apply to us both. The first three chapters deal with the development of a coherent
philosophy of science based on the premise that science must tell us something
about the natural world and the realization that not all science is directed to that
goal. In Chapters IV and V, these criteria are used to force recognition that much
of ecology and limnology, our own work included, is wanting. This revelation
fostered a search for alternative models in limnology (Chapter VI), and ecology
(Chapters VII and VIII) that eventually led to our adoption of empiricism (Chap-
ter IX). The success of an empirical research agenda encouraged us to reassess
the role of an education in science (Chapters X and XI), and provided a sharp tool
in the evaluation of the science, whether in the context of society (Chapter XII),
grant reviews (Chapter XIII), or historical analyses (Chapter XIV). The final
XXIV PREFACES

chapter warns that we must be prepared to change the way we do our science.
If we do not, someone else will change it for us.
There is a need for works that translate between the professional philosophers
and the scientific practitioners in different disciplines, and at different levels of
sophistication. The reader interested in the philosophy of biology already has a
number of choices. Excellent reviews of the history of ecological ideas are avail-
able from Kingsland (1985) and McIntosh (1985). David Hull (1974) and
Michael Ruse (1973) are philosophers who have written widely on the philo-
sophy of biology and biological science. Rolf Sattler (1986) is a biologist whose
book describes the important issues that biology presents to the philosopher. I
even have a rather philosophical book of my own about ecology (Peters 1991a).
These texts offer more sophisticated, more advanced, and usually less ecological
treatments than this book.
Science and Limnology is intended to be an easy read. I have purposely used
less referencing and adopted a less dense style than I would in a scientific paper.
Rather than an authoritative review, it is meant to provide ecologists with a ready
access to the history and philosophy of science. I hope it is sufficiently light read-
ing that both the ecological researcher and the student can find a place for it in
their schedules.
Senior author’s preface
This book is a personal essay expressing the
biases of two researchers and promulgating
their faith in an approach to limnological prob-
lems. As such, the book is little more than a ser-
mon, and warrants consideration only if it is a
reasonably good one. By a sermon, I mean an
essay intended to inspire faith in an, as yet,
undemonstrated and perhaps undemonstrable
property of the universe in which the preachers
implicitly believe.
Science and Limnology is not a monograph that presents new data or develops
a new theory, nor is it a text that reviews the field. The writings collected here do
not fall into one of these conventional categories for scientific works. However, if
the reader has interests beyond normal scientific fare, then this contribution may
be worthy of his or her attention.
Let us assume that the reader’s definition of acceptable reading includes
sermons, and evaluate this book as such. A good sermon has three important
XXVPREFACES
F. H. Rigler

characteristics: (1) It must not be too long; (2) it must be internally consistent;
and (3) it must be comprehensible.
Length: Each chapter is short and the different chapters are sufficiently inde-
pendent, so that only a few pages need be read at a time. Part of the price of this
independence is some disarticulation and a modest amount of repetition among
the chapters. As a result, the book loses some impact and is longer than it needs
to be. Unfortunately, the length of this book was entirely out of my hands, and the
junior author must bear full responsibility if he has made a hash of it.
Consistency: I have fewer qualms on this point. We have both striven to main-
tain a logically consistent position throughout the book. Undoubtedly we have
failed in this intention at many points, but we hope we are no less successful than
many other contributions to the literature. In any case, we knew we were unlikely
to succeed in total consistency before we began.
Comprehensibility: In a sermon, the need to keep the story-line clean is more
significant than length or consistency. The message is very simple. The Baptist
preachers (predictive or empirical limnologists) are trying to persuade the
College of Cardinals (other biologists, ecologists and limnologists) that their
lesser sect has its own valid sources of revelation. Since the Roman Catholics of
science have considered themselves to be omniscient for such a long time, it will
take the patience of Job and the logic and clarity of Thomas Aquinas to convince
them that they can learn from others. We can hardly expect a mass conversion,
but we may help prepare the ground for an eventual reformation.
The intent of the book is to convert those whose belief in traditional ecologi-
cal approaches is weak, and to sow doubts in the minds of those whose belief is
stronger. To do so, we show that some freshwater ecologists have achieved suc-
cess by reflecting on the general nature of science and by developing models that
are consistent with those reflections. We anticipate that a similar approach will
apply equally well to other questions and to other systems. Indeed, a successful
response to the environmental degradation of our planetary home depends on the
widespread adoption of just such an approach. Thus, this sermon has an impor-
tant message. It needs to be heard.
XXVI PREFACES

Prologue
I took advantage of being at the seaside to lay in a store of sucking stones. They were
pebbles but I call them stones. Yes, on this occasion I laid in a considerable store. I dis-
tributed them equally between my four pockets, and sucked them turn and turn about.
This raised a problem which I first solved in the following way. I had say sixteen stones,
four in each of my four pockets, these being the two pockets of my trousers and the two
pockets of my greatcoat. Taking a stone from the right pocket of my greatcoat, and
putting it in my mouth, I replaced it in the right pocket of my greatcoat by a stone from
the right pocket of my trousers, which I replaced by a stone from the left pocket of my
trousers, which I replaced by a stone from the left pocket of my greatcoat, which I
replaced by the stone which was in my mouth, as soon as I had finished sucking it. Thus
there were still four stones in each of my four pockets, but not quite the same stones.
And when the desire to suck took hold of me again, I drew again on the right pocket of
my greatcoat, certain of not taking the same stone as the last time. And while I sucked
it I rearranged the other stones in the way I have just described. And so on. But this
solution did not satisfy me fully. For it did not escape me that, by an extraordinary haz-
ard, the four stones circulating thus might always be the same four. In which case, far
from sucking the sixteen stones turn and turn about, I was really only sucking four,
always the same, turn and turn about. But I shuffled them well in my pockets, before I
began to suck, and again, while I sucked, before transferring them, in the hope of
obtaining a more general circulation of the stones from pocket to pocket. But this was
only a makeshift that could not long content a man like me. So I began to look for some-
thing else. And the first thing I hit upon was that I might do better to transfer the stones
four by four, instead of one by one, that is to say, during the sucking, to take the three
stones remaining in the right pocket of my greatcoat and replace them by the four in the
right pocket of my trousers, and these by the four in the left pocket of my trousers, and
these by the four in the left pocket of my greatcoat, and finally these by the three from
the right pocket of my greatcoat, plus the one, as soon as I had finished sucking it,
which was in my mouth. Yes, it seemed to me at first that by so doing I would arrive at
a better result. But on further reflection I had to change my mind and confess that the
circulation of the stones four by four came to exactly the same thing as their circulation
one by one. For if I was certain of finding each time, in the right pocket of my greatcoat,
four stones totally different from their immediate predecessors, the possibility never-
theless remained of my always chancing on the same stone, within each group of four,
and consequently of my sucking, not the sixteen turn and turn about as I wished, but in
fact four only, always the same, turn and turn about. So I had to seek elsewhere than in
the mode of circulation. For no matter how I caused the stones to circulate, I always
ran the same risk. It was obvious that by increasing the number of my pockets I was

bound to increase my chances of enjoying my stones in the way I planned, that is to say
one after the other until their number was exhausted. Had I had eight pockets, for
example, instead of the four I did have, then even the most diabolical hazard could not
have prevented me from sucking at least eight of my sixteen stones, turn and turn about.
The truth is I should have needed sixteen pockets in order to be quite easy in my mind.
And for a long time I could see no other conclusion than this, that short of having six-
teen pockets, each with its stone, I could never reach the goal I had set myself, short of
an extraordinary hazard. And if at a pinch I could double the number of my pockets,
were it only by dividing each pocket in two, with the help of a few safety-pins let us say,
to quadruple them seemed to be more than I could manage. And I did not feel inclined
to take all that trouble for a half-measure. For I was beginning to lose all sense of mea-
sure, after all this wrestling and wrangling, and to say, All or nothing. And if I was
tempted for an instant to establish a more equitable proportion between my stones and
my pockets, by reducing the former to the number of the latter, it was only for an instant.
For it would have been an admission of defeat. And sitting on the shore, before the sea,
the sixteen stones spread out before my eyes, I gazed at them in anger and perplexity.
[...] And while I gazed thus at my stones, revolving interminable martingales all equally
defective, and crushing handfuls of sand, so that the sand ran through my fingers and
fell back on the strand, yes, while thus I lulled my mind and part of my body, one day
suddenly it dawned on the former, dimly, that I might perhaps achieve my purpose with-
out increasing the number of my pockets, or reducing the number of my stones, but sim-
ply by sacrificing the principle of trim. The meaning of this illumination, which sud-
denly began to sing within me, like a verse of Isaiah, or of Jeremiah, I did not penetrate
at once, and notably the word trim, which I had never met with, in this sense, long
remained obscure. Finally I seemed to grasp that this word trim could not here mean
anything else, anything better, than the distribution of the sixteen stones in four groups
of four, one group in each pocket, and that it was my refusal to consider any distribu-
tion other than this that had vitiated my calculations until then and rendered the prob-
lem literally insoluble. And it was on the basis of this interpretation, whether right or
wrong, that I finally reached a solution, inelegant assuredly, but sound, sound. Now I
am willing to believe, indeed I firmly believe, that other solutions to this problem might
have been found, and indeed may still be found, no less sound, but much more elegant,
than the one I shall now describe, if I can. And I believe too that had I been a little more
insistent, a little more resistant, I could have found them myself. But I was tired, but I
was tired, and I contented myself ingloriously with the first solution that was a solution,
to this problem. But not to go over the heartbreaking stages through which I passed
before I came to it, here it is, in all its hideousness. All (all!) that was necessary was to
put for example, to begin with, six stones in the right pocket of my greatcoat, or supply-
pocket, five in the right pocket of my trousers, and five in the left pocket of my trousers,
that makes the lot, twice five ten plus six sixteen, and none, for none remained, in the
left pocket of my greatcoat, which for the time being remained empty, empty of stones
2PROLOGUE

that is, for its usual contents remained, as well as occasional objects. For where do you
think I hid my vegetable knife, my silver, my horn and the other things that I have not yet
named, perhaps shall never name. Good. Now I can begin to suck. Watch me closely. I
take a stone from the right pocket of my greatcoat, suck it, stop sucking it, put it in the
left pocket of my greatcoat, the one empty (of stones). I take a second stone from the
right pocket of my greatcoat, suck it, put it in the left pocket of my greatcoat. And so on
until the right pocket of my greatcoat is empty (apart from its usual and casual con-
tents) and the six stones I have just sucked, one after the other, are all in the left pocket
of my greatcoat. Pausing then, and concentrating, so as not to make a balls of it, I trans-
fer to the right pocket of my greatcoat, in which there are no stones left, the five stones
in the right pocket of my trousers, which I replace by the five stones in the left pocket of
my trousers, which I replace by the six stones in the left pocket of my greatcoat. At this
stage then the left pocket of my greatcoat is again empty of stones, while the right
pocket of my greatcoat is again supplied, and in the right way, that is to say with other
stones than those I have just sucked. These other stones I then begin to suck, one after
the other, and to transfer as I go along to the left pocket of my greatcoat, being abso-
lutely certain, as far as one can be in an affair of this kind, that I am not sucking the
same stones as a moment before, but others. And when the right pocket of my greatcoat
is again empty (of stones), and the five I have just sucked are all without exception in
the left pocket of my greatcoat, then I proceed to the same redistribution as a moment
before, or a similar redistribution, that is to say I transfer to the right pocket of my
greatcoat, now again available, the five stones in the right pocket of my trousers, which
I replace by the six stones in the left pocket of my trousers, which I replace by the five
stones in the left pocket of my greatcoat. And there I am ready to begin again. Do I have
to go on? No, for it is clear that after the next series, of sucks and transfers, I shall be
back where I started, that is to say with the first six stones back in the supply pocket, the
next five in the right pocket of my stinking old trousers and finally the last five in the left
pocket of same, and my sixteen stones will have been sucked once at least in impecca-
ble succession, not one sucked twice, not one left unsucked. It is true that the next time
I could scarcely hope to suck my stones in the same order as the first time and that the
first, seventh and twelfth for example of the first cycle might very well be the sixth,
eleventh and sixteenth respectively of the second, if the worst came to the worst. But
that was a drawback I could not avoid. And if in the cycles taken together utter confu-
sion was bound to reign, at least within each cycle taken separately I could be easy in
my mind, at least as easy as one can be, in a proceeding of this kind. For in order for
each cycle to be identical, as to the succession of stones in my mouth, and God knows I
had set my heart on it, the only means were numbered stones or sixteen pockets. And
rather than make twelve more pockets or number my stones, I preferred to make the best
of the comparative peace of mind I enjoyed within each cycle taken separately. For it
was not enough to number the stones, but I would have had to remember, every time I
put a stone in my mouth, the number I needed and look for it in my pocket. Which would
3PROLOGUE

have put me off stone for ever, in a very short time. For I would never have been sure of
not making a mistake, unless of course I had kept a kind of register, in which to tick off
the stones one by one, as I sucked them. And of this I believed myself incapable. No, the
only perfect solution would have been the sixteen pockets, symmetrically disposed,
each one with its stone. Then I would have needed neither to number nor to think, but
merely, as I sucked a given stone, to move on the fifteen others, each to the next pocket,
a delicate business admittedly, but within my power, and to call always on the same
pocket when I felt like a suck. This would have freed me from all anxiety, not only within
each cycle taken separately, but also for the sum of all cycles, though they went on for-
ever. But however imperfect my own solution was, I was pleased at having found it all
alone, yes, quite pleased. And if it was perhaps less sound than I had thought in the first
flush of discovery, its inelegance never diminished. And it was above all inelegant in
this, to my mind, that the uneven distribution was painful to me, bodily. It is true that a
kind of equilibrium was reached, at a given moment, in the early stages of each cycle,
namely after the third suck and before the fourth, but it did not last long, and the rest of
the time I felt the weight of the stones dragging me now to one side, now to the other. So
it was something more than a principle I abandoned, when I abandoned the equal dis-
tribution, it was a bodily need. But to suck the stones in the way I have described, not
haphazard, but with method, was also I think a bodily need. Here then were two incom-
patible bodily needs, at loggerheads. Such things happen. But deep down I didn’t give
a tinker’s curse about being off my balance, dragged to the right hand and the left,
backwards and forwards. And deep down it was all the same to me whether I sucked a
different stone each time or always the same stone, until the end of time. For they all
tasted exactly the same. And if I had collected sixteen, it was not in order to ballast
myself in such and such a way, or to suck them turn about, but simply to have a little
store, so as never to be without. But deep down I didn’t give a fiddler’s curse about
being without, when they were all gone they would be all gone, I wouldn’t be any the
worse off, or hardly any. And the solution to which I rallied in the end was to throw
away all the stones but one, which I kept now in one pocket, now in another, and which
of course I soon lost, or threw away or gave away, or swallowed.
Samuel Beckett (1950)
4PROLOGUE

I Why Read about Science?
“If, therefore, a scientific civilization is to be a
good civilization it is necessary that increase in
knowledge should be accompanied by increase
in wisdom. I mean by wisdom a right concep-
tion of the ends of life.”
Bertrand Russell
[The Scientific Outlook (1931)]
Scientists and students are busy. When one of them picks up a paper or a
book, it is with the question, “Why should I read this?” That question is par-
ticularly trenchant for Science and Limnology. This book is not a text or
monograph about freshwater ecology. It is a discourse on the relation of some
of the most fundamental questions in the study of human knowledge to one
scientific sub-discipline, freshwater ecology. It therefore offers few theories
or concepts that busy ecologists might apply in their work or cite in an
upcoming paper. Instead, this book addresses questions that most working
scientists rarely ask: What is science? How does my field fit in? Does my
research belong? What research is worth doing? How can I do it efficiently?
Where should I look for ideas? What ideas are worth teaching? This book is
an idiosyncratic and personal account of my struggles with these issues. It
does not pretend to resolve all the problems, but it does outline how they
affect my science. It offers a starting point for the contemplation of science as
I see it. This chapter establishes the need for such contemplation.
Some Misrepresentations of Science
One indication of the need for such a book is the diversity of views among
biologists in general and ecologists in particular about the nature of science.
Some sense of this confusion can be had from the introductions to general
biology texts. These texts provide an overwhelming amount of up-to-date
biological information; that virtue is not in doubt. Introductory biology books
also allow space to a few paragraphs that purport to define science. These
varied descriptions suggest a flawed grasp of the nature of science (Table 1).

The specific weaknesses of the entries in Table 1 will be clearer when the
nature of science has been explored in subsequent chapters. For the moment,
it is enough to signal the confidence, coherence, and regimentation of these
quotations. They seem to show that science is a straightforward, rational pro-
cess following a set of rules embodied in “the scientific method”. What they
really demonstrate is that biology is a naive and immature science that thinks
too little about what it is doing and where it is going.
Since the writers of biology texts are usually respected scientists, we must
hypothesize that biologists can succeed in science without worrying much
about the general nature of their endeavours. Since less than 1% of their texts
are dedicated to the nature of science, it seems fair to hypothesize that these
leading biologists believe the general nature of science to be far less impor-
tant than the thousands of biological details crowding the remainder of each
6WHY READ ABOUT SCIENCE?
Table 1. Some lapses in descriptions of the nature of science in introductory texts in
biology
The major principle underlying the
experimentation step of scientific en-
quiry is that true hypotheses can never
give rise to a prediction that can be
proved false.
(Weisz and Keogh 1982, p. 9)
An annotated historical appendix, unique
to this textbook among zoology and
biology texts, lists key discoveries in
zoology. (Hickman et al. 1984, p. viii)
The pursuit of scientific knowledge must
be guided by the physical and chemical
laws that govern the state of existence
and interactions of atoms, sub-atomic
particles, molecules and so on.
(Hickman et al. 1984, p. 7)
The ultimate goal of science is to under-
stand the natural world in terms of
concepts, interpretations that take into
account results of many experiments and
observations. These concepts are stated
as theories.... The theory of evolution is
one such conceptual theme.
(Mader 1987, p. 14)
Inductive Reasoning: A logical process
in which a generalization is developed to
explain several specific facts. Hypoth-
eses and theories are formed by inductive
reasoning.
(Brum and McKane 1989, p. 38)
Said briefly, a scientist determines prin-
ciples from observations. This method of
discovering general principles by careful
examination of specific cases is called
inductive reasoning. It first became
important to science in the 1600’s in
Europe, when Francis Bacon, Isaac New-
ton and others began to use the results of
particular experiments they had carried
out to infer general principles about how
the world operates.
(Raven and Johnson 1992, p. 3)
A theory is a hypothesis that has been
repeatedly and extensively tested. It is
supported by all the data that have been
gathered, and helps order and explain
those data.
(Keeton and Gould 1986, p. 4)

text. And since these texts are adopted in hundreds of university courses, we
can further hypothesize that most university professors of biology agree that
the details of their science are far more important than science as a whole.
Confusion about the nature of science is not limited to occasional lapses in
the hurried introductions of first year text-books. A review of the introduc-
tions to theses from research universities and of papers published in leading
ecological journals will provide similar evidence. Too many introductions
justify themselves by pointing at some missing information or uninvestigated
phenomenon that then becomes the goal of the research program. Many biol-
ogists therefore act as though “science” consists of reporting previously
unnoticed facts, and “original research” consists of doing something simply
because it has never been done before. As I will argue below, the appropriate
concern of science is instead the creation, testing and use of theory.
If professionals are little concerned with the general nature of their disci-
pline, it is scarcely surprising that the popular press and electronic media are
also confused about the nature of science. Some echo the professionals —
they see science as a vast search for facts. They write as though all facts are
found in one of two piles (Fig. 1). One, a small heap, consists of all known
facts, and represents the present state of scientific knowledge. The other, a
mountainous pile, represents the facts that science has not yet addressed. The
purpose of science is therefore to move facts from the large pile to the small
one until the large one is exhausted and the small one is immense.
Another popular conception depicts science as a sequence of discoveries.
Newton discovered gravity in his orchard, Darwin discovered natural selec-
tion on the Galapagos, and Fleming discovered penicillin in his lab. In the
same vein, the public hopes and expects that future scientists will dis-
cover cures for cancer, Aids, over-population, and depleted resources, and
7SOME MISREPRESENTATIONS OF SCIENCE
Fig. 1. The erroneous view that science consists of collecting previously unknown facts
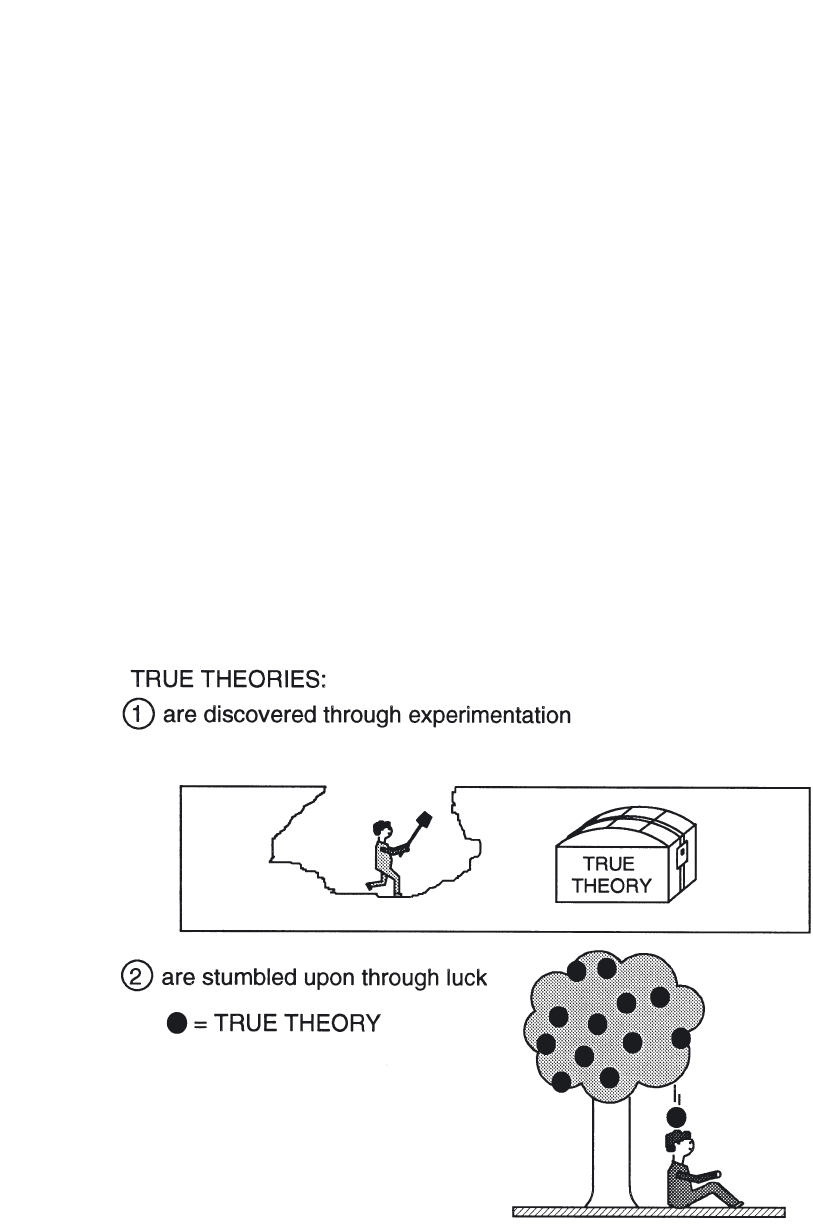
historians can argue whether Priestly or Lavoisier was the discoverer of oxy-
gen. This view sees science as a treasure hunt wherein important principles,
concepts, theories and facts lie hidden by the artifices of stingy Nature. Par-
ticularly lucky scientists stumble across the truth; astute ones wrench truth
from Nature’s unwilling grasp with the right experiment or with a brilliant
intellectual leap (Fig. 2). Others, the unlucky and the dull, overlook vital
clues and wander into ignominy.
An indication that even scientists see science as the discovery of a pre-
existing natural order is provided by the basic protocol of many laboratory
exercises in university teaching. A piece of apparatus is made available to a
student and a question is posed. The student is required to do an experiment
and is prompted to discover the law. The law to be discovered is one posited
by a famous, usually long-dead, scientist. This protocol presents the experi-
ment as the key that unlocks one of nature’s secret boxes. When the box is
opened, the principle pops out and the student makes the one discovery that
was inevitable. If something else is found, the student has failed to repeat the
great scientist’s act of discovery, and (needless to say) the student is wrong.
8WHY READ ABOUT SCIENCE?
Fig. 2. Two versions of the erroneous view
that science consists of discovering the
truth about nature either through good luck
or hard work
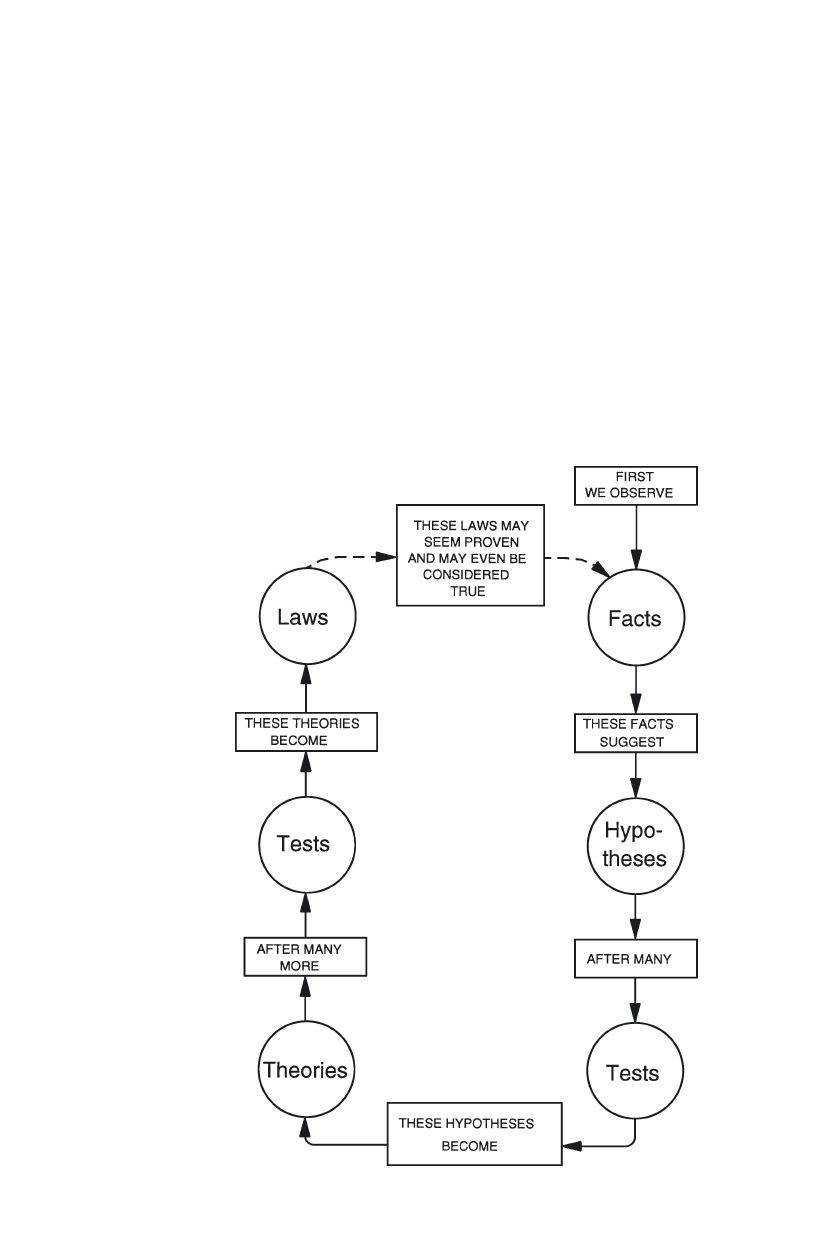
Still another view sees science as the application of the scientific method.
Under this conception (Fig. 3), a scientist begins with a series of observations
or facts, and the careful consideration of these facts results in an hypothesis.
This process is called “induction”. The scientist then deduces other facts from
the hypothesis and “tests” the hypothesis by determining if the deduced facts
are actually observed. If the observations differ from the expectation, the
hypothesis is abandoned. If observation agrees with expectation, the scientist
places a little more faith in the hypothesis and tests it further. As confirma-
tions accumulate, the hypothesis gains in status, becoming first a theory and
finally a natural law.
9SOME MISREPRESENTATIONS OF SCIENCE
Fig. 3. An erroneous version of the scientific method, one that develops true theories
and confuses fact with theory

This description is the treatment that appears in introductory texts. It
seems a serviceable starting point, but it has a number of faults. In practice,
the different elements in the scientific method are much more variable and
much less methodical. For example, scientists often start with an hypothesis
instead of observation; the inductive step may be no more logical than
whimsy; many tests are often biased and flawed; negative evidence likely
exists for every hypothesis, theory and law, but scientists ignore it; most sci-
entists are never aware of a point in time when they made a discovery and
many productive, important researchers are not aware of discovering anything
at all; no theory can be considered “true” or above the threat of disproof in
future tests; and falsification is rarely, if ever, unquestionably complete. To
look at these seeming problems in more detail, I will have to define the basic
elements of the scientific method.
Some Basic Distinctions
Facts and theories. First it is essential that we distinguish between facts
and theories. If we do not do so, we will never understand science. Many
scientists confuse the two. If we look at books or journals, we often see dis-
coveries and laws treated as facts, as suggested in Fig. 3. For example, most
biologists act as if evolution and the elemental table are facts, when they are
really theories.
A fact is an observation, a datum or sense impression which has not yet
been consciously interpreted and about which no scientific claim is being
made. By itself, a fact is empty and useless, because it gives no basis for
action. Parenthetically, one may object that such a pure fact probably cannot
exist, that all observations are interpreted through our biological and cultural
biases as soon as they are sensed. In other words, all facts are “theory-laden”.
A stricter interpretation would define a fact so that, as soon as it became more
than empty and useless, the fact would be termed a theory. I accept this, but
will not complicate discussion further for the time being.
A theory is a generalization that goes beyond the facts. It therefore makes
predictions which are statements about facts we do not yet know. Consider a
simple example. If I were to rise in a scientific meeting to state “it is not a fact
that animals need food to grow”, most of the audience would dismiss me as a
lunatic. Nevertheless, it is not a fact that animals need food to grow. It is far
more important. It is a theory. The facts are that the effect of starvation of a
few individuals of a few species has been studied. Each of these individuals,
when deprived of food, failed to grow, and instead wasted away and died. We
do not know that other individuals of the same species would do the same. We
10 WHY READ ABOUT SCIENCE?

can never know what every individual of every species that ever lived would
do. Therefore, we are going far beyond the facts when we claim that all ani-
mals need food to grow. When we state a theory, we make a statement that we
can never show to be true. We can however demonstrate that a theory is prob-
ably false as we shall see in the next section.
Induction and deduction. Deduction is the derivation of specific in-
stances of a generalization. In simpler terms, deduction is the process by
which we decide what predictions a theory makes. For example, suppose that
we are interested in the growth of babies, that we measured and weighed a lot
of them (Table 2) and that we had erected a theory (Fig. 4, overleaf) to
describe the relationship between the weight (in kg) and length (in cm) of
babies:
Weight = 0.0002(Length)
3
(1)
Perhaps few would be willing to say we have made a discovery, but by
developing an equation relating length and weight, we have created a theory,
albeit a very simple and trivial one. There are other theories that would
describe the data just as well; some of those may be much more comprehen-
sive than Eq. (1), and they may offer deeper explanations, but I will postpone
discussion of those complications. It is sufficient for the reader to accept that
one characteristic of a scientific theory is that a theory offers the power to pre-
dict about specific cases on the basis of a more general statement. Most
authorities concede the point.
The equation is a theory for two reasons. First we did not measure all
babies, so we are claiming that we can extrapolate from our observations to
babies we have never seen. Second we did not measure all sizes, even of the
babies we did measure; so we are claiming that we can interpolate the weights
that these babies had or will have at lengths we have not observed. When we
go beyond the facts by extrapolating or interpolating, we have made a theory.
11SOME BASIC DISTINCTIONS
Table 2. Some facts about the lengths and weights of some babies
Baby Length (cm) Weight (kg)
Vic 39 11.864
Sarah 30 5.400
Eva 25 3.125
Adrienne 32 6.554
Mike 29 4.878
Julian 22 2.130
Elisa 37 10.131
The theory in Fig. 4 makes some very definite predictions. It says that for
any baby we measure, there will be one, and only one, possible weight, that
predicted by Eq. (1). This weight is a deduction from the theory. It is the log-
ical consequence or implication of the generalization or theory. The impor-
tant point is that a given theory makes specific predictions about unobserved
instances, and that anyone who uses the theory will make exactly the same
predictions. The theory may be wrong, because not all babies behave as it
suggests, but that would not affect the claim that the relation is a theory.
Deductions allow us to test the theory to see if the predictions are correct.
Given a theory, like Eq. (1), and some additional specific cases that were not
used to build the theory, say Anna or Kate, we can make the required mea-
surements of the babies’ lengths, and calculate or deduce their expected
weights. We can then test these predictions by observing the babies’ weights
and comparing them to our expectations. If Anna or Kate is lighter or heavier
than we predicted, we would have falsified our theory and therefore we would
have to revise it.
In describing the creation of a theory, I skipped over one of the most con-
tentious parts, induction. Induction is the process by which we move from a
set of facts to a theory or generalization. This is where intuition or inspiration
enters science. The scientist looks at the facts and they, in some mysterious
way called induction, suggest a generality to the scientist. Induction is not a
logical process. It does not lead inevitably to one conclusion, but instead
could lead to an infinity of possible theories.
12 WHY READ ABOUT SCIENCE?
Fig. 4. A theory about the size of babies
The point that the same set of facts could inspire different researchers to
entirely different theories seems hard for many people, including some scien-
tists, to accept. I will therefore develop a ridiculously simple illustration
(Fig. 5). The example will again deal with growing babies but this time, just
for variety, we shall develop a theory about how fast babies grow. We will
again begin by making a few measurements of weight (W, in kg) and age (X,
in months), and after one month’s work we might find:
Age Weight
11
22
Now I need only sit back, contemplate the data and wait for an inspiration.
After some time, I might hit on a more general pattern and propose a theory
to describe the growth of a baby:
W = X (2)
13SOME BASIC DISTINCTIONS
Fig. 5. The development of alternative theories about the effect of age on the weight of
babies

A more sophisticated, or at least more mathematically competent, ob-
server might instead propose another observation on the basis of the same
observations:
W = e
0.346X
(3)
In fact there is an infinite series of relations, each representing a theoreti-
cal generalization, that could describe these data.
Logic will never tell us which of these simple theories best describes the
growth of a baby, but we can logically deduce the consequences of the two
competing theories. If Eq. (2) is correct, then W = 3 kg when X = 3 months.
But if Eq. (3) is correct then W = 4 kg when X = 3 months. Assuming that we
still have our baby, we need only wait until it is 3 months old to take the
observation that will allow us to determine which theory makes the better pre-
diction. If we find that W = 4 at X = 3, we can reject W = X as the less useful
theory. We thus know that W = X is wrong, but we cannot therefore claim that
W = e
0.346X
is correct. Someone else might look at the three pairs of data
points and induce that:
W
X
= X + W
X–1
(4)
where the subscripts indicate weight at the ages of X and X–1 months.
Since Eq. (4) fits the available data as well as Eq. (3), we cannot tell which
of these two competitors, or of a host of other possible theories connecting
the points, is right or wrong or even better. However, once again we can
deduce the logical consequences of the competing theories. When X = 4,
Eq. (3) suggests that W
4
= 8, but Eq. (4) would instead suggest that W
4
= 7.
Once again, we must wait a month and make more observations to test these
alternatives. Obviously, we will never reach the end of this process. We could
postulate various wavy lines or step functions implying varying growth rates
within each month, or an infinity of other possibilities that could join the four
weights of the growing baby. In fact, no matter how many points there are,
there will still be an infinite number of relations that could connect them
together, and because we always use induction to arrive at the theory to
describe these points, there can never be any guarantee that we have found the
right one. In other words, we can never know if we have found the truth and
therefore we should never claim that a theory is true in the sense that it is
beyond doubt.
This is a profound discovery. It is one that Aristotle never made, Galileo
rejected, and Newton dismissed. The widespread acceptance of the principle
that we can never be sure of anything in science is one of the most revolu-
tionary changes of the 20th century.
14 WHY READ ABOUT SCIENCE?

Empirical and explanatory theories. The theories I have used to illus-
trate this discussion are modest statements of trend that describe some data
and could be used to predict future observations of the same kind. Many sci-
entists are unwilling to call these statements “theories”. I see this as only a
problem of definition.
Like many who have considered the problem, I call any statements that
make predictions “theories”. This single term embraces two general types
which I call “empirical theories” and “explanatory theories”. Empirical theo-
ries are simple relationships based on observed patterns (e.g. allometric
relations, species-area curves, the second law of thermodynamics) that
make predictions by assuming that the future relations among the variables
will resemble those in the past. Explanatory theories are grander, more
comprehensive statements that not only make predictions, but also offer a
comprehensible explanation of the phenomenon being predicted, like the
size-efficiency hypothesis of Brooks and Dodson (1965), the trophic cascades
of Carpenter et al. (1985), Darwin’s theory of evolution by natural selection,
and various theories of creation, like the big-bang hypothesis. Both empirical
and explanatory theories play important roles in science.
The history of scientific achievement shows that theories develop in a
characteristic sequence. First the scientist surveys the subject matter of inter-
est, looking for patterns that might allow a prediction. This might be the
swirling of stars in the night sky, or the occurrence of rain, or the production
of certain chemicals in the lab. This search seems a simple step but it is just
the opposite. The patterns we are seeking are invariably set among a vast col-
lection of confusing and unrelated facts. These irrelevant observations often
obscure the regularity of the system under study.
Once the scientist has perceived a regularity in the objects of study, and
expressed this pattern unambiguously, an empirical theory has been pro-
duced. Empirical theories may be complex, but they are more likely to be
simple X-Y relationships because such relations are so easy to perceive and to
define. Thus the relations describing the weight of a baby from its age or
length are theories. They are theories because they generalize from our obser-
vations and allow the prediction of weight from any value of age or length.
Empirical theories can be useful, but they have limitations. First, they
make predictions only about the correlated variables. Thus, they can never
generate unexpected predictions about other aspects of these entities or about
other phenomena. They remain theories of relatively low generality. Second,
they do not satisfy our desire to explain our environment; this sense of under-
standing seems to be a bodily need, at least for some researchers.
Empirical theories stimulate other scientists to explain why the theories
work. With luck, someone will eventually replace the original empirical
15SOME BASIC DISTINCTIONS

theory with an explanatory theory. Because this new theory relates the
observed variables to other observations, it may make more predictions and
thus be more useful. Given the limited predictive power of empirical theories,
many scientists underplay their importance in science. Nevertheless, the his-
tory of scientific development shows that empirical theories are a crucial step
toward explanation.
In summary, science is not about collecting facts or observations. It is the
process of identifying, testing and organizing generalizations or theories that
go beyond the observed facts and, in some sense, explain these facts as
instances of a general pattern. Both empirical and explanatory theories are
created by induction, an inspired guess that leads us to postulate some regu-
larity in the universe. They are then tested by deduction that allows us to
know what the theory predicts so we may compare its predictions to further
observations.
The Importance of Science
Why should we care what science is? The reasons are clear. As citizens,
we expect science to give us some grasp of the risks that we face and some
measure of control over our fate. As taxpayers, we ultimately fund research,
and so we must learn to scrutinize the growing demands of modern science
for money and resources. As teachers and students, we need to know what
science is so that we can teach and learn what is scientifically important, and
disregard what is trivial. As scientists, we need a better knowledge of what we
are doing, so we can use our resources well to confront the problems of sci-
ence and humanity, so we can promote our vision to the rest of society, and so
we can defend it from unjust criticisms. We cannot afford to be ignorant of the
nature of science any longer. Society must learn what it can expect from sci-
ence, and what it cannot.
There seems little call for a detailed defense of the thesis that science
plays an important role in our civilization and therefore that the nature of
science should be better appreciated. Like it or not, our food, shelter, health,
our ways of life depend on science and technology. We may deplore this
dependency as unnatural, dangerous, and unstable, but we cannot change the
situation. Science has helped create the problems of the 20th century, but
to throw away science at this stage in civilization would be a fatal error.
We now depend on science so much that the solution to our dilemma will be
scientific, whether it is civilized or catastrophic. We must use science for
good, because we must use science. That will be easier if we know what
science is.
16 WHY READ ABOUT SCIENCE?

Many humanists have taken the trouble to discover what science is. They
often do a better job of explaining science than do the scientists. It may rankle
scientists to turn to humanists to discover the nature of science. However,
there is no reason to disdain their help.
The prologue to this book is an allegory of scientific research. It shows
that the novelist and playwright, Samuel Beckett, had a keener appreciation
of the nature of scientific enquiry than many scientists. There is science in
Molloy’s struggles with the problem of the sucking stones, in his abandon-
ment of the vaguely felt, yet fundamental principle of trim, and in the
inelegant (but sound) solution whose elaboration marked the end of both
enquiry and interest. In all of this, Molloy gives a better picture of science
than most introductory biology texts.
Jacques Barzun is another humanist who has concerned himself with sci-
ence. His Science: The Glorious Entertainment is a milestone. In addressing
C. P. Snow’s (1963) concept of two cultures, Barzun writes about the need to
appreciate science:
The new science is for the public a Delphic mystery; it keeps the western
intellect troubled but unenlightened, except for the practitioners them-
selves. Here we touch upon the grim deficiency of the scientific culture,
which is also the first lesson to be drawn from our historical review: the
fundamental lack in our mental and spiritual lives does not come from
the trifling division between scientists and humanists, or between scien-
tists and the whole of the laity; it comes from the fact that science and the
results of science are not with us an object of contemplation.
By common consent the sciences are taught in school and college;
but everyone admits that this teaching is wasted on three fourths of those
who are forced to endure it. For all the use or interest they find in “the
science requirement” they might as well be required to take Greek or
Latin: science remains to them a dead language. The belief persists that
if sciences were better taught they might prove to be more titillating, but
there is little agreement on how to teach them. And one reason for this is
that to the teachers and practising scientists themselves science is rarely
an object of contemplation. They teach therefore as if to intending
professionals. (Barzun 1964)
Barzun’s condemnation is well deserved. Somehow in a society that
depends on science, we manage to turn off most students. How do we do it?
We teach as we were taught, and we were taught “as to intending profession-
als.” Given a choice, few professors would elect to teach any students other
than those concentrating in biology, and most of us prefer to teach the details
of our own narrow specialities. If instead we want to instruct a larger fraction
17THE IMPORTANCE OF SCIENCE

of the general population about science, we must escape the traditions of our
own training. We must find ways to make science meaningful to more people,
without necessarily recruiting them to the profession. We must teach science
as an object worthy of contemplation. I feel our only hope of doing so is to
understand something about the nature of science. We scientists must step
back from the welter of detail that bemuses us in our daily work and learn
from enlightened humanists. If we can do so, we may be able to talk across
the gulf that sometimes separates humanist and scientist, and across those
that separate us from our students, our public patrons and our colleagues.
The Growth of Science
Derek de Solla Price (1986) dedicated much of his career to measuring the
size of science. He showed that whether one measures science by the number
of scientists engaged, the number of papers produced, the number of journals
published or the number of dollars spent, one reaches the same conclusion:
for the past 300 years, science has doubled in size every 10 to 20 years. In
other words, science doubles about twice as fast as population (Fig. 6). This
has some remarkable implications. One is that over 85% of all the scientists
who ever lived are still alive. Another is that the scientific literature increases
18 WHY READ ABOUT SCIENCE?
Fig. 6. A comparison of the growth rates of science and population in the United States.
(Modified from Price 1986)

by about ten times over the course of an individual’s career and has done so
since its inception; we have always been awash with new work and we always
will be. A third implication is that science has grown by encroaching on the
rest of society.
The growth of science has demanded accommodation from working sci-
entists. For example, we assemble in multi-disciplinary teams of specialists to
teach and do research, we insist that scientists write, and that students read,
even longer text-books, and we use the power of the computer and other
machines in our data collection, data analysis, writing and publication. Many
of us work nights, weekends, and holidays. We may focus our efforts more,
and hone our interests to narrower specialties. Some ruthlessly excise erst-
while non-professional interests from their lives — hobbies, friends, family.
Many scientists I know have already done these things, but we have not
resolved our difficulties. We may only have made things worse. Science is
growing faster than we can handle.
Obviously, science cannot grow faster than society forever. For example,
if science does not slow down, every man, woman, and child in North Amer-
ica will have a Ph.D. by the year 2200. Whom, or what, will we teach then?
Research now consumes 2 to 4% of the gross national product of most devel-
oped countries. If science were to continue to grow at present rates, this will
increase to between 4 and 8% in another generation, and between 16 and 32%
in another 100 years. There must be an upper limit and the explosive growth
of science must eventually slow as that limit is approached. Since science
already commands a large share of resources, we should expect to see the
reduction in growth in the near future.
A slower rate of growth will present new problems, and require some hard
decisions. For example, it is unlikely that society will fund all future mega-
projects of all the disciplines, or meet all the demands of all the researchers.
Eventually, choices will be made among many seemingly incomparable
proposals. For example, we may have to decide if science is better served by
a space station, a biotechnology complex, a mass accelerator, or an oceano-
graphic vessel. We may have to compare the desirability of research into
biodiversity with that into chemical pollution or crop yields. We may have to
decide among sub-atomic physics, molecular biology, astronomy and limnol-
ogy. To do that, we will likely have to know what science is, not just what
interests its sub-disciplines.
Summary
I can now return to the question in the title of the chapter: Why read about
science? First, a growing literature shows that science can be appreciated and
19THE GROWTH OF SCIENCE

understood as a process, rather than simply as a body of information, so that
we can expect our reading to be profitable. Second, there are many indica-
tions that science is not properly appreciated. Introductory texts and graduate
theses show that many biologists are unsure of the nature of science and the
role of the scientist; I believe that an appreciation of science would make us
better teachers and produce better students. There is also evidence that the
fundamental components of research — facts, theories, tests, induction and
deduction — are poorly understood by many members of society; a better
grasp of these basics would allow the citizen to function better in a society
that funds and depends on scientific research. It may also allow scientists to
do their jobs better. Finally, we should learn what science is and what it is not,
because the dynamics of growth of humanity and science imply that the
traditional relations between science and society will soon change. Modern
researchers and citizens are about to experience interesting times, and I
believe that an appreciation of the nature of science will help them find a way
out of the gathering confusion.
20 WHY READ ABOUT SCIENCE?

II A Brief History of Method
“The great scientists ... represent to me a simple but
impressive idea of science.... This, then, for me is
science. I do not try to define it, for very good rea-
sons. I only wish to draw a simple picture of the
kind of men I have in mind, and of their activities.
And the picture will be an oversimplification.... My
criterion of demarcation between science and non-
science is a simple logical analysis of this picture.”
K. R. Popper
[The Problem of Demarcation (1934)]
Much of this book is premised on Karl Popper’s distinction between scientific
theory and non-scientific concept, his criterion of demarcation. According to
Popper a construct qualifies as a scientific theory if it makes a potentially
falsifiable statement or set of statements. These statements are called “pre-
dictions”. The main goals of science are to make theories, to use theories to
make predictions and to assess those predictions against observation.
Although Popper’s views are widely known, they are not universally
accepted even by philosophers, and their relevance is even less appreciated by
many working scientists. For example, I was raised in a tradition that science
discovers the truth, that only a scientist can understand science, and that
philosophers and historians had nothing to tell the researcher. I came to real-
ize the folly of that position through a consideration of the history of biology,
a history that often reflects the stages of my own philosophical development
and often those of other scientists. This chapter therefore uses an historical
overview of biological thought as a device to appreciate Popper’s criterion of
demarcation.
Three Ways to Knowledge
For all the variety of human thought and endeavour, there are only three
methods of gaining information about the universe: the intuitive method, the
metaphysical method, and the scientific method. Each method is character-

ized by certain assumptions about nature and by criteria for judging insights
that the methods produce.
The intuitive method is the oldest way of discovery. It assumes that no
laws govern the behaviour of natural objects or, if there are laws, they can be
repealed at any time by some greater organizing being or force. Given the
fickleness of observable phenomena, the intuitive method puts little faith in
observation. Instead, it holds that the only way to gain knowledge of lasting
value is through revelation. Truth will be revealed to us if we put ourselves in
so receptive a frame of mind that the organizing force can reveal its inten-
tions. It is a corollary of this view that humans, by their own efforts, cannot
gain any real understanding of the universe or hope to predict future events.
The second method, the metaphysical method, has been favoured by many
philosophers. It assumes that the behaviour of natural objects is governed by
a set of laws and that people can discover these laws by their own efforts.
This method emphasizes the power of the human mind to see these laws. It
begins with the search for premises that are obviously true and, by the use of
logic, deduces natural laws from these axiomatic truths. Both the intuitive
and metaphysical methods justify their insights by interpreting appropriate
observations as instances of the perceived general truths.
Finally, the scientific method shares the metaphysicians’ assumptions that
the universe functions according to laws. Scientists differ from metaphysi-
cians in that they rely not only on thought, but also on observation to define
these laws. Science may therefore begin with observations, as suggested in
Fig. 3, or with an idea. Wherever the method begins, scientists use their
brains to produce a theory or law. Then they move to test the implications of
theory against new observations. If the theory fails to predict well, the theory
is changed and the new theory is tested in its turn.
The essential difference between the scientific method and the others is
that the statements of science need not be truths. Science does not require its
practitioners to perceive truth directly, nor does it accept that others can do
so. In science, knowledge is always hypothetical, so the ultimate arbiter in
science should be the testimony of observation, not the fervour of belief.
The three methods of enquiry are not necessarily applied separately. Time
has allowed hybridization, and the history of science can be very usefully
interpreted as the history of the interplay of these three methods.
Aristotle
We begin with Aristotle (384–322
BC) because science arguably began
with the Greeks and Aristotle was the most influential of the Greek scientists.
22 A BRIEF HISTORY OF METHOD

We are only interested in his method of finding out about nature. When we
look at how Aristotle did that, we find a curious blend of metaphysics and
science.
Aristotle should be considered a scientist because he insisted on the
importance of observations. For example, 2300 years ago it was commonly
believed that the human embryo nourished itself by grasping the mother’s
flesh with its mouth and sucking. Aristotle ridiculed this idea because “any-
one who takes the trouble to look” will see that the embryo is completely
enclosed in a fluid filled bag; it could not possibly suck the mother’s flesh
(Bodenheimer 1953). Aristotle concluded that nourishment must enter via
the placenta and umbilical cord (Fig. 7A, overleaf). This is what we still
believe today.
Aristotle’s argument for placental nutrition, although it concerned a small
point, was a piece of science. He built on one theory — that all animals need
nutrition — and one observation — that the fetus is enclosed in the amniotic
sac — to conclude that the prevailing theory was false. He then drew on a fur-
ther observation — that the umbilical cord passed from mother to fetus — to
produce a generalization that concerned all human embryos: the embryo is
nourished through the placenta and cord.
In considering the problem of placental nutrition, Aristotle appears to
have thought very much as scientists do today. However, elsewhere in his
writings, we encounter arguments that are almost meaningless to us. At those
points, Aristotle blends science with his now strange metaphysics. A com-
mon property of all these now meaningless arguments is that they derive from
“intelligible principles”.
Aristotle’s intelligible principles were really general statements, or if you
wish theories, that seemed to be self-evidently true or axiomatic. They are
like the obvious truths that characterize the metaphysical method of studying
nature. For example, for Aristotle it was an intelligible principle (a) that the
universe is perfect, and on a less general scale (b) that the circle is the perfect
geometrical form. A consequence of these principles is that we should find
circles everywhere. Aristotle did (Fig. 7B). He saw instances of these princi-
ples in the paths of the planets, in the cycle of birth, life and death and, since
evolution or extinction would disrupt this perfect cycle, in the permanence of
the species. Given this bias, Aristotle would not look for extinction or evolu-
tion of living species, and would probably have disregarded any observations
relevant to those processes. As a result, he used his principles to deduce that
species are permanent.
The argument for placental nutrition seems essentially scientific even
today, but that for the permanence of the species is almost ludicrously incom-
prehensible. I have described these two instances of Aristotle’s science to
23ARISTOTLE
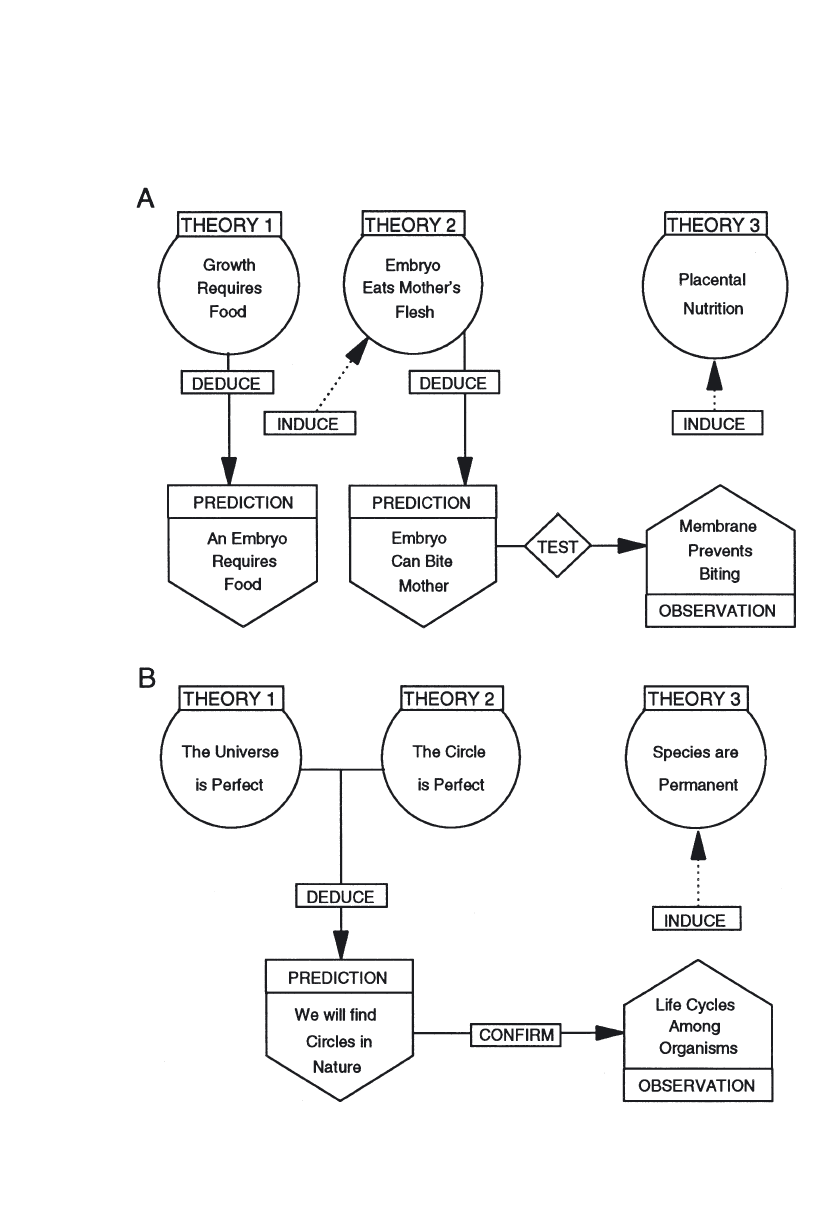
24 A BRIEF HISTORY OF METHOD
Fig. 7. (A) Aristotle’s use of the scientific method in developing the theory of placental
nutrition. (B) Aristotle’s use of the metaphysical method in arriving at the theory of the
permanence of the species

contrast the metaphysical and scientific methods of enquiry Aristotle used to
understand the universe.
In both cases, Aristotle was confronted by theories of uncertain origin.
Some, like the theory that the embryo sucked the mother’s flesh, were pre-
sumably common-sense beliefs of the time. Others, like the theories that the
universe and the circle are perfect, arose from his mind as intelligible princi-
ples. We cannot reject a theory because of its origins. We insist that the the-
ory make predictions, but predictions can be deduced from both theories in
Fig. 7. Both methods moved Aristotle to make other observations and to
develop other theories, like those regarding placental nutrition and the per-
manence of the species.
The difference between the scientific and metaphysical methods lies in the
way different practitioners link prediction to observation. Aristotle the scien-
tist looked for observations that could be inconsistent with prediction. When
they were inconsistent, he rejected the theory as false and moved to create a
better theory. Aristotle the metaphysician looked for observations that were
consistent with the theory because he wanted to illustrate the truth of his
intelligible principles. Because the principles were not under test, he saw con-
trary evidence not as falsifying but as irrelevant. Since the intelligible princi-
ples were not open to change, the induction of the permanence of the species
was not intended to be tested, but to extend the whole complex of theory and
observation so that as much “knowledge” as possible would be consistent
with the intelligible principles.
This explains why Aristotle’s work is alternately meaningful and mean-
ingless to us. When guided by intelligible principles, his work seems crazy,
because it reflects the world view of a brilliant man who lived in an almost
unimaginably different time and culture. Whenever he is free of these princi-
ples, whenever he saw no connection between the theories and the principles,
his use of observations becomes “scientific” and valuable.
Deduction and Induction in the Age of Reason
Throughout the dark ages, Aristotle’s ideas, beliefs and observations were
accepted almost without question. In a sense, both his observational science
and his metaphysics were treated as indubitable principles, so the distinction
between the two methods of enquiry became blurred. When science began to
emerge in the 16th century, there was a strong reaction against this meta-
physical science. This reaction went in two diametrically opposed directions.
One faction is associated with Francis Bacon (1561–1626), an English
philosopher and visionary, who also served as Lord Chancellor to James I.
25DEDUCTION AND INDUCTION IN THE AGE OF REASON

Bacon recommended that we first purge our mind of wild theories and intel-
ligible principles, and then carefully and systematically collect a mass of
observations relevant to the subject of interest. From this material, Bacon
thought we would induce a general theory, or two or three theories that could
explain the observations. He supposed that we would then disprove and elim-
inate the wrong theories by careful observations and experiments. Bacon’s
method was characterized by its absence of intelligible principles and its
emphasis on observation and induction. Subsequently, the Baconian method
has been associated with the less tenable opinion that induction proceeds log-
ically and certainly from masses of systematic observation to truth, but that is
a perversion of his more complex views (Eiseley 1973).
Another approach was taken by the French philosopher and mathemati-
cian, René Descartes (1596–1650). Descartes was impressed by the certainty
of mathematics, which advances step-by-step from one indisputable conclu-
sion to another. Any claim to true knowledge outside of mathematics seemed
dubious. He therefore concluded that we must bring order into chaos by
applying the clear and certain deductive method of mathematics to philoso-
phy and science.
Deductive science would only be possible if the deductions started from
one or more very general and true theories. Descartes saw that the intelligible
principles of Aristotle and his medieval followers had led only to confusion,
and so devoted himself to a search for indisputable axioms that could form
the basis for a true science. By rigorous thought, he arrived at two proposi-
tions he considered indubitable: I think, therefore I am; and God exists.
Descartes’ radicalism got rid of a lot of the spurious truths that had directed
the science of Aristotle and his successors. However, Descartes also left us a
one-sided view, still dependent on intelligible principles and still limited to
deduction.
Both Descartes and Bacon greatly influenced subsequent views of science.
Two schools of thought developed around their teachings. One tells us that
knowledge is obtained by induction and the other relies on deduction. The
resulting methodological debate eventually led to the newer and much greater
question of whether it is possible to obtain true knowledge in any way at all.
This terrible question undercut a basic assumption about the quality of
knowledge and the goals of understanding. This was a turning point for
science and humanity.
David Hume and Immanuel Kant both sought to answer this new question.
In doing so, they came to very different answers. Each was extremely influ-
ential and each founded a philosophical tradition based on his answer.
David Hume (1711–1776) was a Scottish philosopher and writer. Interest-
ingly (and somehow reassuringly) he thought his major philosophical treatise
26 A BRIEF HISTORY OF METHOD

was a failure that “fell stillborn from the press” (Popper and Miller 1983).
Before Hume, mathematics was considered a science, so analogies between
the two were used to support the belief that science yields true knowledge.
Hume instead separated the two, and so made that argument for the truth of
science untenable. Hume then showed that induction did not give us certain
knowledge.
I can only approximate the reasoning that led him to this belief. Philoso-
phers differ about his arguments and I have no reason to suppose that I can do
better than the professionals. Hume started with the demonstration that we
know nothing about the external world. All we have are sensory impressions.
Therefore we do not know that events in the external world regularly follow
one another. We only know that certain sensory impressions follow each
other in time.
For example, if a sequence of two events, P then Q, happens in the exter-
nal world, we can never know it. All we experience, all we know, is a succes-
sion of sensory impressions, p then q. From these impressions, we infer the
occurrence of the corresponding sequence of real events, P then Q. However
this inference is a guess or induction. Because it is not certain, we can never
know that the real events exist. We all sometimes experience this uncertainty
when we become confused about whether a memory occurred in reality or in
a dream. If we cannot know what events are real, we can similarly never
know if there is a causal connection between P and Q. If there is no causal
connection, then we can never be sure that Q or q will follow P or p. No
matter how many instances of p then q we experience, we can never claim
that we are sure that the next p will be followed by q. We have even less
reason to believe that Q will necessarily follow P.
Hume concluded that the only systems about which we have true knowl-
edge are artificial, like mathematics, and that we can never know when we
have true knowledge about the universe. In the terms I have been using, there
is no logical basis for believing that induction will ever lead us to a true
theory about nature.
The great German philosopher Immanuel Kant (1724–1804) led the clas-
sical backlash to Hume. Kant argued that our knowledge of the external world
does not merely come from Hume’s sensory perceptions. For Kant, knowl-
edge has two sources. There is empirical or a posteriori knowledge that
comes after the fact, through our senses, as Hume agrees. But there is also a
priori knowledge which is supplied by our consciousness independently of
all experience. Of these two, a priori knowledge is by far the more important
source.
In his Critique of Pure Reason, Kant argued that the most basic laws of
nature can be discovered a priori, that is before the fact. Thus, discovery does
27DEDUCTION AND INDUCTION IN THE AGE OF REASON

not require science or empirical observation, it could be achieved by pure
reasoning alone. In fact, Kant held that if we did not have these a priori laws
for perception to play upon, we could not have any experience of the world at
all.
With Hume and Kant, there appears to be a clear schism. Hume argues that
the only source of knowledge is empirical but that it is not reliable, whereas
Kant, Descartes and Aristotle found that reason is the most reliable, and per-
haps the only, source of knowledge. Philosophers have not resolved this
dichotomy, but natural scientists have tended to follow Hume. His denial of
the possibility of certain knowledge was at first devastating, unacceptable,
and even incomprehensible, but if scientists had instead followed Kant, they
might have had to abandon observation and then they could scarcely claim to
be natural scientists.
Logical Positivism
The philosophical direction that developed from Hume reached one ex-
treme in the writings of Ernst Mach and his associates. These philosophers
were known as the Vienna circle and their philosophy subsequently became
known as logical positivism. Their history and ideas are explored by Phillipp
Frank in his Modern Science and its Philosophy (1949) and his The Philo-
sophy of Science (1957) upon which I depended in writing this section. At one
time, I was deeply influenced by their writings. I no longer believe that they
represent an end-point to philosophical development, because their philo-
sophy is so extreme as to be completely unacceptable for many scientists.
However, some of the findings of logical positivism still address contempo-
rary issues in biology and so serve to show how thinking about science can
help scientists to do their work. Just as importantly, the extremity of logical
positivism prepares us to accept the more recent and useful position of Sir
Karl Popper.
Hume viewed observations as only sensory impressions and held that the-
ories suggested by these observations need not bear any relation to the laws
of nature. Logical positivism suggested the situation might be far worse:
Maybe there are no natural laws at all, for how would we know if there are?
The regularities we perceive, and the explanatory theories we build around
them, may be nothing more than a product of the scientists’ minds. Perhaps
the human mind demands the existence of laws, and scientists work in such a
way as to satisfy that need. If so, the laws of nature may be no more than a
definition of how scientists’ minds work.
On causality. As an example of this argument, I will develop Phillipp
Frank’s attack on causality. Many scientists see causality as the cement of
28 A BRIEF HISTORY OF METHOD

the universe; they hold causal connection among events to be the basis for
meaningful pattern in the universe and contend that the description of these
connections is the goal of science and the basis of theory. Frank attacks this
fundamental assumption because, if he can topple that, the rest of our self-
assurance about science and reality should follow.
Frank tried to show that the law of causality only exists because scientists
act in such a way as to preserve it. In everyday words, we could express this
law as “every event or effect has a specific cause”. Frank states the law more
rigorously: “If a State A of the universe is once followed by a State B, then
whenever A occurs again, it will be followed by B”. This pure form of the law
can find no application in science because we can never know the state of the
whole universe. But if we do not know the state of the whole universe, we can
never be sure that some additional factor will not modify the State A so that
State C follows instead. Frank feels that what we do is to reformulate the law
as a more useful approximation: “If in a finite region of space, State A is once
followed by State B and at another time by State C, we can make State B as
similar to State C as we wish by increasing the size of the region of space we
consider”. This reformulation is more meaningful if we translate it into the
everyday language of experiment: “If State A once appeared to be followed
by State B and another time by State C, then our results were spoiled by an
unknown and uncontrolled variable”. To paraphrase Frank further, when we
apply the law of causality to a finite biological system, the number of vari-
ables we control and the number of organisms we sample are determined by
the amount of agreement we demand among our replicates, that is by how
similar we require B and C to be.
Once he showed that the law of causality is rigorously valid only for an
infinite system, Frank proceeds to a second argument. Even if we suppose the
law to be valid for small, finite systems, there is still reason to doubt that it is
really a natural law. Science begins with data and these data are sensory
impressions. Our statements and generalizations must always refer to some-
thing we sense. We must be able to see them, touch them, smell them or
something. Our laws and theories must therefore be translated, or at least
translatable, into terms that have meaning in relation to what we observe.
Whatever version of the law of causality we choose, the law refers to the state
of the system. Frank defines this state in sensory terms as the sum of all per-
ceptible properties of the system. However, when we find that some observa-
tion apparently violates the law, we invent unperceived, even imaginary,
properties of the system to explain the contradiction.
Frank provides an example. Imagine two pairs of identical iron rods on a
table. Both pairs are in State A. One pair simply sits there, that is State B. The
other pair move towards each other — State C. To satisfy the law of causal-
29LOGICAL POSITIVISM

ity, we say that the latter pair of rods had an imperceptible property called
magnetism. Thus we conclude that the initial states were only apparently the
same, and in doing so we preserve the law of causality. If we act this way, the
law of causality will always be obeyed because whenever a system does not
obey the law we invent as many fictitious properties as necessary to preserve
the law.
Frank concludes that the law of causality no longer looks like a law. It is
simply a definition. It defines the way we will interpret any situation in which
A is not inevitably followed by B. If we treat causality as a law, we merely
introduce a modern intelligible principle to science.
On new ideas. Frank’s ideas on causality seem radical. After all, we have
used the concept of magnetism since we were children, whereas discussion of
States A, B, C and causality is unfamiliar and highly philosophical. Many
biologists might be tempted to dismiss logical positivism as empty intellectu-
alism. If this is so, an examination of Frank’s views about how we convince
ourselves of the truth of intelligible principles and general theories is appro-
priate.
His thesis is that a theory, when first introduced, is incomprehensible. We
do not understand it and do not use it unconsciously as a premise in our think-
ing. Therefore, when a theory is new, we treat it as a theory should be treated,
as a construct that may be “scientifically valid, but philosophically false”. It
is scientifically valid in that it makes more and better predictions than com-
peting theories, and it is philosophically false because we do not believe it
represents an eternal and self-evident truth. As we become more accustomed
to the theory we begin to have more faith in it. Finally, we come to take the
generality for truth.
Frank illustrates his point by reviewing how thinkers’ attitudes to New-
ton’s laws changed with time. Shortly after Newton published his theory of
gravitation, the Irish philosopher and cleric George Berkeley (1685–1753)
objected to it. He felt that scientists take a lot of trouble to apply, use and test
their theories, but do not try hard enough to understand them. How can a the-
ory be valid if we do not understand it? Berkeley saw that Newton’s laws
made good predictions, but found them incomprehensible. They were scien-
tifically valid, but philosophically false.
The German philosopher and scientist Gottfried von Leibniz (1646–1716)
also objected to Newton’s theory. He wondered how a moving body could
keep its direction and velocity with respect to empty space, and how bodies
can exert a force on one another through empty space. For Leibniz, both
experience and Aristotelian physics tell us that these elements of Newton’s
theory were in conflict with our perception of reality. For Berkeley and
Leibniz, Newton’s theory was philosophically false because it offended both
30 A BRIEF HISTORY OF METHOD

reason and common sense. However, it was scientifically valid. Conse-
quently, it was used by scientists and technologists for generations. It still is.
With the passage of time, Newton’s theory was built into the thoughts of
philosophers, scientists and educated laymen. It became assimilated and by
the late 18th century, it had become philosophically true. For example,
Immanuel Kant claimed to show that the law of inertia could be derived from
pure reason and believed that, unless we accept the law of inertia as true, we
will never understand nature.
In a few generations, Newton’s laws changed from being philosophically
false to being philosophically true. They eventually became a necessity of
thought. This internalization was so complete that when Einstein showed that
Newton’s laws were scientifically incomplete, and even false, it was a stun-
ning blow to the concept of scientific truth.
Sir Karl Popper
Logical positivism leaves us with the conclusion that science is not about
the real world at all. It is a description of the way that scientists’ minds orga-
nize sequences of sense impressions. As a philosophy of life and work, I do
not find this very attractive. It removes many misconceptions, but offers little
that one can use to improve one’s science. However, it is a powerful device to
free us of our own intelligible principles and to prepare us accept a different
philosophy. Logical positivism helped us escape the view that science dis-
covers the whole truth about nature. In doing so, it prepared the way for the
views of Sir Karl Popper.
Popper was born in 1902 and trained as a physicist. In Vienna, he came to
know the members of the Vienna circle and their ideas. His most influential
work was The Logic of Scientific Discovery, first published in German in
1930 and in expanded form in English in 1959. He moved to England to
escape Nazism and has since published many books and papers in English.
Good summaries of the vast corpus of his works are available in reviews by
Magee (1973) and Pera (1980), and in an anthology of his writings edited by
Miller (1985).
Popper’s contribution was to give scientists a beautifully simple scheme
for recognizing and evaluating science. He accepts much of logical posi-
tivism, including the conclusion that a theory can never be shown to be true.
Knowledge therefore cannot be a true understanding of natural laws; it is only
the ability to look into the unknown to predict as yet unobserved events.
These predictions are achieved by constructs that we call theories. Because
science is the activity that generates such theories, science is the only source
of such knowledge. Most importantly, the only good theory is one that has the
31SIR KARL POPPER

potential to be wrong. If no conceivable observation could show the theory
wrong, then the theory must predict every possibility and therefore would tell
us nothing. In other words, to qualify as a scientific theory, a statement or set
of statements must be potentially falsifiable. For example, the statement that
human babies will weigh less than 15 kg could be wrong, so it is a theory. The
statement that human babies will weigh something is not.
For Popper, scientists begin by creating a theory. It is irrelevant where this
theory came from: principle, observation, revelation, intuition, induction, or
simply a lucky guess. What is important is whether the theory identifies any
potential future observations or facts as inconsistent with the theory, because
the theory makes a prediction by telling us what facts will not be observed.
We can then test the theory by comparing future observations with those
facts. If unexpected observations are made, the theory has been falsified. If
the facts agree with the predictions, we would be justified in using it again.
Popper’s work has given us a clear working description of theories and
facts. The former are the devices we use to make predictions and the latter are
the instance that the theories predict. This great clarification should allow us
to communicate better with both scientists and non-scientists, and to do better
science. Because it is a scheme that deals only with the relation between
theory and observation, and not with intelligible principles or philosophical
axioms, we can use it, regardless of future philosophical changes. Indeed,
much of the philosophical debate that has subsequently focused on Popper’s
work seems irrelevant to me as a working scientist. Popper has freed us from
the philosophical fashions that make so much of Aristotle and other philoso-
phers incomprehensible.
For me, Popper’s work was a revelation. I was troubled by Hume’s argu-
ment that induction was fallible and verification impossible, and could see
no escape from the answer of logical positivism, that science was simply the
creation of our minds to describe their own workings. I was not satisfied with
science or philosophy.
Popper accepted the reality of the philosophical difficulties that I could not
escape, but he also saw that these things did not matter. Science works regard-
less. What makes it work is its method and central to that method is
falsification. As a working scientist I need no longer worry about being
wrong, because it is irrelevant to ask if my theory is a true representation of
the external world. I no longer need to ask if this theory is more true than that
theory. I need only ask:
(1) Does it predict anything?
(2) Does it predict more than its rivals?
(3) Can I show that the theory is wrong?
32 A BRIEF HISTORY OF METHOD

Furthermore, I can do my science knowing that I have profited from a long
philosophical tradition and that I base my thoughts on a current philosophy
whose fundamental characteristics are not likely to change. Contemplation of
the nature of science has made me a happier and better scientist.
33SIR KARL POPPER


III Normal Science and Pseudo-Science
“It is the normal practise of scientists to ignore evidence
which appears incompatible with the accepted system of
scientific knowledge, in the hope it will eventually prove
false or irrelevant. The wise neglect of such evidence pre-
vents scientific laboratories from being plunged forever
into a turmoil of incoherent and futile efforts to verify false
allegations.”
Michael Polanyi
[Personal Knowledge (1958)]
Popper’s approach has many virtues, but his science is an abstract ideal.
When Popper illustrates his ideas, he uses heroic figures like Newton and
Einstein. Popper therefore gives a goal to strive for, but not a picture of what
most real scientists do. His prescriptive definition of science must be tem-
pered with a descriptive one that reflects better the reality of science as a
human activity. Thomas Kuhn has provided one such description.
Real scientists, those who do day-to-day science, are also real people.
They want recognition, they feel the pressures of society, and they suffer var-
ious limitations of intellect, temperament, resources, and opportunity. Such
people probably comprise 99.9% of all scientists who ever lived, and this
chapter examines their work. It begins with a review of the masterful work of
Kuhn on the nature of everyday science, and ends by considering the recep-
tion of challenges to the scientific community, in light of Kuhn’s ideas. This
book, like many contemporary essays in the field, uses the word “science” in
both Popper’s prescriptive and Kuhn’s descriptive senses, but the context
should make my intention clear.
Kuhn’s “Normal” Science
In 1962, Thomas Kuhn published what might be one of the most important
books about science in this century, and certainly one that has had a continu-
ing influence on me since I first read it in 1968. The Structure of Scientific
Revolutions continues the process of uprooting our faith in science as an

austerely intellectual search for greater truth about more phenomena. But
instead of a reanalysis of the philosophy behind the scientific enterprise,
Kuhn offers an historical reanalysis.
Many histories of science treat past research either as unsuccessful
attempts to reach, or as necessary intermediary steps to achieve, contempo-
rary ideas. Kuhn rejected that approach. Instead, he provides a new and
unflattering model about what scientists actually do and have done.
An historical model of science. Kuhn posits that any branch of science
cycles between long periods of “normal science” when scientists follow a
dominant tradition or “paradigm” and short periods of crisis and revolution-
ary change when the paradigm is shaken and ultimately replaced. Kuhn also
postulates an early pre-paradigmatic stage before the discipline emerges.
That phase seems an appropriate beginning for a review of his model (Fig. 8).
Pre-paradigmatic science. The earliest members of a scientific discipline
are at a considerable disadvantage, because the discipline cannot yet be well
defined. There are no text-books to describe the material of the discipline, no
courses to provide model answers to model questions, no scientific societies
to identify the proud tradition that contemporary members of the discipline
should respect and advance, no manuals of methods, nor any reviews of
important questions.
The founders of a field are free of both the guidance and the constraints of
tradition. As a result, there is usually a great deal of confusion and flounder-
36 NORMAL SCIENCE AND PSEUDO-SCIENCE
Fig. 8. A schematic representation of Kuhn’s model of scientific revolutions

ing in the fledgling discipline. Terminology is inconsistent, methods are
suspect and even the relevance of various observations is questionable. More
positively, the field is very open because there is no constraining tradition.
Different schools of thought compete and intellectual ferment is real. In the
absence of specialists, researchers are drawn from many existing disciplines.
To talk to one another, they must disseminate their new ideas in accessible
language, and since contributions cannot assume a common intellectual base,
books are an honoured device for communication. Relevant observations are
often so simple, that the new discipline is even open to educated laymen.
Normal science. Eventually, researchers reach some modicum of agree-
ment on the nature of the field. They embrace some view about how their cor-
ner of the universe works, at least in general terms, and this view dictates
reasonable avenues of investigation. Working within this world-view, prac-
titioners develop standards for education, research, and reporting. They come
to accept certain questions as significant, and to ignore others as irrelevant.
They produce intellectual leaders, text-books, courses, and learned societies,
and these promote specific theories, applications, instrumentation, and
methods.
In Kuhn’s terminology, these common scientific traditions represent a
paradigm or series of paradigms for the new field. Paradigms are what we
refer to when we speak of Copernican astronomy, or Newtonian physics, or
evolutionary biology. These paradigms not only tell scientists what to expect
of the world, they also define legitimate subjects for future research and the
methods for that research. In Kuhn’s view, textbooks serve to propagate the
paradigms of the field, and the purpose of scientific training is to indoctrinate
a new generation of scientists to think and act like their teachers. Once the
paradigm is so established that the practitioners in the field have a well
defined research program, the field may be said to enter the phase of “normal
science”.
Before Kuhn, the accepted view was that science advances as a struggle
between competing theories (Platt 1964). A winning theory is consistent with
all relevant facts, whereas the loser is not. Kuhn instead argues that contrary
evidence is available for every theory at all times, but that scientists unite
behind their favourite anyway. Normal science therefore engenders agree-
ment among researchers, not confrontation. One might wonder why science
does not grind to a stop. The answer to this question sets Kuhn apart from
other observers of science.
Kuhn suggests that an emerging paradigm is very limited in scope and
precision. It is often more successful than alternatives only in solving those
problems that a group of scientists have recognized as particularly acute. It
out-competes the opposition and becomes established, not necessarily
37KUHN’S “NORMAL” SCIENCE

because it solves more problems, but because it promises to solve more. The
allure for scientists is that the new theory provides more questions for them to
address.
Kuhn’s normal science is a mop-up operation as scientists attempt to real-
ize the promise of their paradigm. “Normal scientists” are those who try to
squeeze nature into the conceptual mould provided by their paradigm.
Because no theory is perfect, they must often ignore, dismiss or simply not
see anomalous observations. Thus normal scientists often seem to be striving
to confirm their theories, not to test them. Kuhn suggests that even great
figures act as normal scientists for most of their careers, and only rarely as
revolutionaries. He believes that good scientists must always maintain an
“essential tension” between conservatism and radicalism (Kuhn 1977). As a
result, normal science is the business that occupies most scientists for most of
their lives.
Kuhn identifies three classes of activity that typify normal science. Some-
times researchers who seem to be testing the theory are really looking for
confirmatory instances and trying to show how useful the paradigm is. Other
times they may be observing certain phenomena more carefully and precisely
than ever before, because the paradigm suggested that such efforts may lead
to new discoveries. For example, after Newton, more exact measurements of
the position of the stars and planets allowed the discovery of other heavenly
bodies; after Dalton, more exact estimates of atomic weights allowed the
discovery of isotopes; and after Darwin, careful study of heredity led to
genetic theory. The third activity in normal science seeks to resolve the
theory’s initial ambiguities and failings.
According to Kuhn, normal scientists do not revolutionize the field, and
likely are never famous. Kuhn’s answer to the obvious question about why
they do their work is disturbing, yet it echoes Barzun’s view of science as
entertainment. Kuhn’s normal scientists do their work because they enjoy
puzzle-solving. They have a question which is certified as valid by the
paradigm, they have a set of rules for solution that the paradigm has also
specified, and they have a large part of the solution that the paradigm has
again supplied. The motivation for science is the same as that for art, history,
music and all the other activities that keep humanity occupied: our own dis-
interested curiosity, perhaps the one truly human activity.
Crisis and revolution. Kuhn’s normal science sows the seeds of its own
destruction. Eventually normal science ceases to provide us with interesting
enough puzzles, so some scientists point to previously ignored anomalies as
an indication that the old paradigm is in crisis and begin to look elsewhere for
gratification. During this period of uncertainty, the anomalies may provide
fodder for research within the old paradigm, so some workers rearticulate
38 NORMAL SCIENCE AND PSEUDO-SCIENCE

theory and practice to account for the now troublesome anomalies. Neverthe-
less, as more scientists become interested in observations that do not fit, the
old paradigm begins to fragment. An alternative approach or paradigm even-
tually promises more fertile ground for future research. The old paradigm is
abandoned, the new one takes its place, and a new generation of normal
scientists begin the cycle again.
Kuhn ends with the question of scientific progress. In a Kuhnian scientific
revolution, some problems that the old paradigm addressed may be dropped
from consideration as uninteresting. Kuhn interpreted these shifts in interest
as indications that different paradigms are incommensurate. Thus successive
paradigms cannot be compared fairly, and there is no basis to claim that new
paradigms are better than the old ones.
Kuhn’s model makes science a very human subject, and very different from
the intellectual discipline of the philosophers. His science is only “what sci-
entists do”, and is much closer to our everyday use of the word than Popper’s
heroic examples. Nevertheless, the differences between Kuhn and Popper are
smaller than they may seem (Kuhn 1977). In particular, Kuhn’s model is con-
sistent with the philosopher’s conclusion that we cannot recognize truth in sci-
ence. Indeed, given the scepticism of modern scientific philosophy, we prob-
ably feel less that we understand nature than did most of our predecessors.
Criticisms of Kuhn’s model. Despite the richness and even genius of
Kuhn’s book, some aspects of his model seem less sound. Although it is not
my intent to review his position fully, I cannot leave the topic without point-
ing to some shortcomings.
The model implies that all sciences pass through a pre-paradigmatic stage.
However, such a phase would be very difficult to identify in any area of
human interest. Theories and paradigms existed in most areas long before the
emergence of science. For example, before there was an evolutionary biology
or a uniformitarian geology, there was a creationist paradigm, and there
seems to have been some such paradigm since the dawn of humanity. Perhaps
a pre-paradigm phase is only an illusion that one sees in retrospect.
Another criticism focuses on Kuhn’s use of the scientific papers of normal
scientists as evidence for his thesis. These papers often contain confirmatory
tests of dominant paradigms. The authors’ words suggest that they are
pleased that their tests support the paradigm and that they did their
work to protect that paradigm. However, when researchers write a paper, the
rationale may have to be rephrased to fit the results. The introductions there-
fore present an idealized and flattering picture of scientific foresight, because
any other introduction would confuse the purpose of the paper and confound
the reader. If a scientist wanted desperately to overturn a paradigm but failed,
the finished paper would almost certainly describe the work as a successful
39KUHN’S “NORMAL” SCIENCE

test of the dominant ideas in the field (Medawar 1990). Kuhn may have over-
interpreted these politic misrepresentations as indicators of unthinking sup-
port for the dominant paradigm among scientists.
My final criticism is more general. Kuhn’s book is short and readable, but
this was achieved in part by excluding examples. Tests for his ideas are there-
fore difficult to devise because he provides so few models. Indeed, when one
tries to identify a paradigm, a crisis, a pre-paradigm stage, or even a field of
normal science, Kuhn’s intuitively attractive concepts prove surprisingly
difficult to apply. We discover that paradigms exists within paradigms, that
hierarchies of normality, crises and revolutions make any identification of
the current phase of a science ambiguous. Seemingly consistent episodes are
therefore easy to find because the model is slippery and the observations
malleable. For similar reasons, critical tests are rare (Cohen 1985).
Kuhn’s model is therefore not without problems. Perhaps that is appropri-
ate for a potential paradigm of the nature of science. After all, initial prob-
lems are what the model predicts.
“Pseudo-Science”
Kuhn’s analysis helps resolve one of the continuing challenges to scien-
tific researchers. It explains why most of us dedicate our lives to the resolu-
tion of apparently minor details of nature, rather than addressing glamorous
questions of broad interest to society. Why do we not leave our laboratory
benches to study alien abductions, holistic medicine, extra-sensory percep-
tion, or spectral apparitions? Kuhn’s answer is that our scientific tradition
identifies research on these topics as inappropriate. Such material can be
called “pseudo-science”, even if it passes Popper’s criterion of demarcation,
because it falls outside the Kuhnian model whereby science is largely what
normal scientists do.
In this sense, pseudo-science is anything that purports to be science, but
that the established scientific community does not accept as such. So defined,
pseudo-science is not the delusional domain of a lunatic fringe. For example,
Gregor Mendel spoke to the Brünn Natural History Society on two occasions
in 1865 about his experiments with inheritance in plants; he published this
work the next year and informed the leading scientist Karl von Naegli about
the importance of the work in 1867. Nevertheless, as far as the world of sci-
ence was concerned, Gregor Mendel did not exist. No one paid any attention
to his work until 1900 when it was rediscovered simultaneously by three
botanists (De Vries, Tschermak, and Correns) and Mendel became the father
of Genetics.
40 NORMAL SCIENCE AND PSEUDO-SCIENCE

What I am suggesting is that from 1865 to 1900, Mendel’s work was
pseudo-science, because it was treated as pseudo-science. For 35 years no one
recognized it as science. It was simply ignored. There is nothing unusual
about the treatment Mendel received. Most new ideas about science are
ignored. No one gets excited about them, no one cites them, they just disap-
pear unnoticed. That is the fate of most pseudo-science.
A further example demonstrates the other extreme of the scientific
response to pseudo-science. In 1910, the German meteorologist Alfred
Wegener became convinced that, 200 million years ago, all the continents
were joined into a single super-continent he called Pangaea. Wegener pro-
moted this theory for many years and it excited considerable debate amongst
geologists until 1929. That year, at the Geological Congress, Wegener’s
theory was officially declared to be false, and so became pseudo-science for
the geologists. Curiously, there was widespread, if tentative, acceptance of
Wegener’s views among biologists, even after it had been rejected by geolo-
gists; so a theory could be pseudo-science under one tradition while it was
still science under another. There may be no right or wrong in science, but a
very similar distinction is maintained between what the majority of scientists
declare as interesting or uninteresting. That is the point of these examples of
pseudo-science.
The year after his theory was declared false, Wegener himself disappeared
in Greenland where he had gone to test his theory. The theory did not die. A
number of new discoveries, like the lack of sediment in parts of the ocean
floor and the mirror images of fossil magnetism on either side of the mid-
oceanic ridges, eventually forced a complete re-appraisal. Today, the ‘new’
theory of plate tectonics is universally accepted.
This cruel and unfair treatment of new hypotheses is just what one should
expect. According to Kuhn, we scientists spend most of our careers trying to
patch up our leaky theories, modifying and mending them in the hope that
they will one day fit the facts. As long as the theory holds the promise of
being patchable, as long as it continues to provide interesting puzzles to keep
us amused, there is no chance that the majority of scientists will reject it. We
will cling to the established paradigm as tenaciously as a child clings to a
favourite toy.
We will not reject a favourite theory simply because someone shows us a
few embarrassing facts. Neither will we accept a new theory simply because
some rebel champions it. There is a time for new theory, and that time is when
our old theory has ceased to entertain us by generating new, but soluble,
puzzles. This forces us to look to the anomalous observations that we had pre-
viously ignored as pseudo-science. Thus Mendel was a pseudo-scientist in
1865, but by 1900 his time had come. We were ready for a new toy.
41“PSEUDO-SCIENCE”

The reverse of this reaction to a new theory is also part of the reception of
pseudo-science. As long as we are content with the dominant paradigm, the
theory it represents moves gradually towards acceptance as a universal truth.
As such, the paradigm tells us what sort of experiments we can do, what
methods we should use, what canons of evidence we will accept, and what
form of publication we should use.
The initial reaction to the theories of Mendel or Wegener could be
discounted as near-sighted conservatism that was eventually put right by
forward thinkers. In that case, the examples would show the adequacy of
scientific self-correction, despite the relativism of scientific truth and the
dominance of the paradigm. This alternative explanation seems to posit that,
although science was always wrong in the past (because no past theory has
survived unchanged), modern science is right. I prefer the relativistic expla-
nation, not just because it avoids hubris, but because it renders the scientific
conservatism of the past more rational and comprehensible. The conserva-
tives were not benighted disciples of error, but conscientious scientists work-
ing under other premises than those we use today. The lesson we should learn
from Mendel and Wegener is to be more tentative in our conclusions about the
validity of scientific theories.
Velikovsky. My last example of pseudo-science has not enjoyed the re-
habilitation of Mendel or Wegener. It has always been considered pseudo-
science and seems likely to remain so. My purpose in introducing such mate-
rial is not to champion a failed cause, but to illustrate the resistance of science
to new paradigms, the advantages and disadvantages of such resistance, and
some of the mistakes that characterize a failed scientific revolution.
Immanuel Velikovsky was born in Russia in 1895. He studied abroad, but
returned to Moscow to take a medical degree and subsequently practised
medicine and psychiatry in Tel Aviv. After 15 years, he began a study of
Akhenaton, Oedipus and Moses, during which similarities in the legends of
different civilizations convinced him that a global catastrophe took place
about 1450
BC. While seeking the cause of such an event, he discovered that
neither Babylonian nor Hindu astrologers ever made mention of Venus
among the planets, even though it is one of the brightest objects in the sky. He
concluded that Venus did not exist during early human history.
The theory. From these and many more observations, all derived from
ancient texts, Velikovsky developed the following theory and published it
in Worlds in Collision (Velikovsky 1950). Our solar system originally had
one less planet than it does now. Sometime before 1500
BC, a violent explo-
sion on Jupiter gave birth to a massive comet that eventually became the
planet Venus. About 1450
BC, this comet passed close to the earth, producing
enormous tides, global heating, electrical discharges and a rain of hydrocar-
42 NORMAL SCIENCE AND PSEUDO-SCIENCE

bons. These events are associated with the exodus of the Jews from Egypt.
The comet then disappeared into space, only to return around 747
BC, when it
collided with Mars. This collision brought the comet into planetary orbit as
Venus but so perturbed the orbit of Mars that in 687
BC, Mars almost collided
with Earth, causing one or more new global catastrophes. Although they may
not have been so devastating as the upheaval of 1450
BC, they were enough to
tilt the earth’s axis by 10°.
According to Velikovsky, this theory explains many myths and legends,
explains the absence of Venus from early astronomical records, and explains
some of earth’s petroleum deposits (as hydrocarbons from the planet’s tail).
He also believed that the theory predicted that (a) Venus would be hot, (b) its
atmosphere would contain hydrocarbons, and (c) Jupiter would emit radio
waves. Neither these predictions nor others are very clearly stated in the
book. Nevertheless, Velikovsky believed that he had made these and other
predictions, and that all of them proved correct (Anon. 1972).
Velikovsky followed Worlds in Collision with a second book, Earth in
Upheaval (Velikovsky 1955), in which he gathered together a large number of
biological and geological observations that he believed were consistent with
his theory. He places before the reader a whole series of both well-known and
obscure facts about vast bone heaps in the Arctic, about whale skeletons on
hilltops, about erratic boulders found hundreds of kilometres away from their
parent bedrock, about the food in the mouths of quick-frozen mammoths
in Siberia, etc. These observations serve three purposes. First, Velikovsky
wanted to convince us that many observations do not fit existing geological
theory. Second, he wants us to accept that there was a recent world-wide cata-
clysm, and to change the date of the last ice age so that it will be consistent
with the hypothesized approach of Venus in 1450
BC. Finally he wants to
convince us that biological evolution could not have taken place by mutation
and natural selection as Darwin supposed, but that it arose through mass
extinctions caused by the cataclysms, followed by rapid speciation through
multiple mutations caused by heat and radiation associated with the cata-
clysms.
The reception of Velikovsky’s theory. At the time, Velikovsky appeared
even more radical than he does now. His theories about rapid speciation pre-
dated the contemporary interest in discontinuous rates of speciation called
“punctuated equilibrium” (Eldredge and Gould 1972) by a generation. His
belief that single celestial events might control major turning points in terres-
trial natural history predated current theories about mass extinction and mete-
orite impacts (e.g. Melosh et al. 1990) by 30 years. And his explanation of
events in human history as the result of dramatic changes in their non-human
environment was equally far ahead of its time.
43“PSEUDO-SCIENCE”

Velikovsky managed to challenge the fundamental paradigms of four
fields simultaneously. Not surprisingly, he was not well received. Most scien-
tists simply and totally ignored his work. Some wrote damning criticisms of
his book. Five of these critics had never read it. Other scientists brought such
pressure to bear on his publishers (MacMillan) that they transferred the rights
to the book to Doubleday and Company which was less easy to pressure
because it published no textbooks. The unseemly condemnation of the author
and his ideas by the scientific and academic establishment (de Grazia et al.
1966, Rose 1972, Stove 1972) made Velikovsky something of a hero during
the 1960’s. His book sales made him a millionaire, and eventually he was
appointed to the Princeton Institute for Advanced Studies, where Einstein
had worked. But he never found scientific respectability for himself or his
theories.
Why did Velikovsky’s theory fail to win support? If we accept Thomas
Kuhn’s view of science, it is clear that Velikovsky made a number of fatal
mistakes. He failed to present scientifically acceptable evidence, he failed to
present his work in the proper form, and he tried to revolutionize fields that
were not ready for change.
Velikovsky’s data sources were ancient texts, including the Bible. For him,
observations like the parting of the Red Sea by Moses and the fall of manna
on which the Israelites fed were anomalous facts that the dominant paradigm
could not explain. For most scientists, these stories were simply irrelevant;
they were not facts at all. Most scientists saw the history of their discipline
partly as a liberation from such ideas. Scientists did not cite holy books; they
used the theories of physics, the concepts of mathematics, and the resolving
power of fine spectroscopes and powerful telescopes. Velikovsky’s sources
seemed a long step backwards. A serious work of science would be packed
with tables, graphs and equations, but Velikovsky wrote 400 pages without
any. By the standards of science, but not his own, Velikovsky presented no
data at all.
Velikovsky also failed to examine any other explanation for the observa-
tions he presented. Although we scientists cannot hope to be unbiased
towards our intellectual creations, we are expected to vet these ideas as care-
fully as possible by considering alternatives. Velikovsky passed this onus to
the reader and thus sacrificed his own credibility.
Velikovsky also used the wrong style to present his ideas. Compared to
the dry, impersonal writing of the professional researcher, his prose looks
like sensational journalism. Moreover, Velikovsky chose to publish in popu-
lar books, rather than in scientific journals. Scientists have developed formal
standards for communication, the scientific paper and the monograph (Kinne
1988), and are deeply suspicious of those who ignore those standards.
44 NORMAL SCIENCE AND PSEUDO-SCIENCE

Not only did Velikovsky fail to present his theory effectively, he also
chose to present it at the wrong time. According to Kuhn, a new theory can
only be effective when the old paradigm has entered a state of crisis, when it
has ceased to provide a bounteous supply of soluble puzzles. Unfortunately
for Velikovsky, his books appeared when traditional sciences had more puz-
zles than they had scientists to solve them. Astronomy was still ecstatic about
the giant telescope at Mount Palomar, and radio-telescopes were in the offing;
the theory of stellar evolution was still fresh, the nuclear furnace was new,
and the consequences of the red shift were being explored. Geologists were
also delighting in a surfeit of new tools: deep-drilling for stratigraphy, mag-
netometry, seismographic surveys, and soon satellite pictures and remote
sensing; intellectually, they dealt with the evolution of rock types and the
discovery of new minerals, and were on the brink of the new theory of plate
tectonics, a theory which would eventually address many of Velikovsky’s
objections. Biology was even healthier. Chromatography and the electron
microscope soon led to the development of molecular biology; evolutionists
were pursuing the possibilities of the new synthesis between genetics, natural
history, selection and molecular biology. Scientists felt no need for a radical
new paradigm, when normal science offered so much.
Finally, and perhaps most importantly, Velikovsky failed to give a clear
and unambiguous description of his theory. He hints at its beauty, like a
dancer behind seven veils, but the veils are never lowered. The theory is
never apprehended. By any scientific standard, such vagueness is unaccept-
able.
Testing Velikovsky’s theory. Kuhn’s model explains the lack of interest in
Velikovsky’s theory, but not why the theory was ridiculed rather than being
subjected to scientific tests. I therefore raise two further questions: “Can
Velikovsky’s theory be falsified?” And if so, “Why was it not tested?”
I believe that Velikovsky’s theory can be tested and that if it had been
tested, it would have been falsified. In a discussion of tree-rings in relation to
the catastrophism, Velikovsky tries to convince the reader that no trees sur-
vived the cataclysm of 1450
BC and that there had been violent climatic
changes in 747 and 687
BC. To achieve his first objective, he describes the dat-
ing of the giant sequoias of California, and concludes that “the most ancient
of these started life after the year 1300 before the present era”. This is
advanced as evidence that no tree survived the “great catastrophe of the
middle of the second millennium”. He dismissed from consideration the
oldest tree of all — the General Sherman — on the grounds that it had not
been cut down. However, everyone knows that you can age a tree without
cutting it down. In fact, the General Sherman tree had been aged in 1946 and
its germination date estimated as 1550
BC ± 500 years. Thus there is a
45“PSEUDO-SCIENCE”

good chance that this tree antedated Velikovsky’s great cataclysm. Since
Velikovsky published, even better evidence against the great cataclysm has
been found. A number of bristlecone pines have been aged and appear to have
begun life between 2050 and 2950
BC.
A second category of tree-ring data can be used to test Velikovsky’s
theory. Annual growth varies with climate, so dramatic changes in the width
of the annular rings might indicate climate change. Velikovsky found evi-
dence for such changes that might represent the cataclysms of 747
BC and
687
BC (Douglas 1919), but failed to mention that even greater changes
occurred many times in the life of these trees. Furthermore, he neglected to
mention that very large annual fluctuations in tree growth are correlated with
very slight differences in mean temperature and precipitation in the recent
record. Thus we must conclude that there is no evidence for any major distur-
bances over the entire period from 750 to 660
BC. Similar evidence for the
absence of cataclysms can be had from the record left in lake sediments.
If Velikovsky’s theory can be falsified with observations like this, one
must wonder why it was not done. I cannot speak for others. For myself I did
not bother to publish these tests or to devise others because these ideas
seemed so futile. Why would anyone want to disprove something that no one
believes (at least, no one whose scientific opinion is worth having)? If some-
one claimed that Mars was made of Edam cheese, what joy would scientists
have in disproving the theory? What place would they win in the estimation
of their peers or in the history of science? As Polanyi suggested, the normal
practice of scientists is “wise neglect”.
46 NORMAL SCIENCE AND PSEUDO-SCIENCE

IV The Ecologists’ Disease: Two Personal Examples
“In dealing with any aspect of limnology, as perhaps
any other branch of science, it is impossible to avoid
the thought that no work is perfect and that the
greater proportion of published investigations are
very imperfect indeed. Every one of us is at fault in
some way or another, every one of us must attempt to
achieve progressively higher standards in accuracy,
scope and imagination.”
G. E. Hutchinson
[The Prospect Before Us (1966)]
Ecologists have known for over a generation that something is wrong with the
earth. There are too many people, too few wild places, too much waste, too
little clean water, too many chemicals, too little food. These troubles make
the life of an ecologist interesting, if depressing. We have been dragged from
the quiet of academia to help solve the global problems of over-exploitation
and over-population.
We have tried to respond effectively. Everywhere, universities and col-
leges have produced courses, programmes, chairs, institutes and faculties to
deal with “human biology” and “environmental science”. Some ecologists
spend their evenings in legal challenges to proposed developments, political
movements, and preaching to the public. Ecological researchers offer a dizzy-
ing array of new techniques, concepts and instruments to address our prob-
lems. A suite of learned societies have been formed and a host of new journals
are being published. This is an exciting time because society desperately
needs the services of good ecologists, drawn from the full range of science
and beyond.
Nevertheless, for many ecologists, ecology has not provided the quest we
sought. We entered the science because we felt a love for nature and the
beauty of natural things, and because these things are under threat. Then, as
soon as we were trapped in our careers, teachers confronted us with theoreti-
cal models, multivariate statistics, simulations and hypothetico-deductive
frameworks. Our colleagues express horrified disbelief when we admit

ignorance of eigenvalues, matrix algebra or Laplace transforms, and we
naturalists hide our inadequacies like guilty secrets. It sometimes seems that
we can only save the things we love by abandoning them in our work.
Even as we accept our inadequacy, we make another horrifying discovery.
Different experts are equally confident, but they all have different ideas. At
this point, we should feel bewildered.
My own bewilderment first caused me pain, but then it led me to wonder
how much of what we call ecology can be considered scientific. Other ecolo-
gists are also questioning the relevance of ecological research, asking if their
work is useful, and if they could be more effective. These questions spring
from a deep disillusionment with the present paradigms of ecology. I am
encouraged by such questions, because the present confusion is a sign that
better times may be in store for us. I think ecology is ready for a Kuhnian sci-
entific revolution (Kuhn 1962) and I see criticism as the goad that will push
us to change (Peters 1991a).
I contend that the body of ecology is infected by a strange disease. The
infection is not new, but its extent was unappreciated until our troubles were
brought into the open by a set of unusual circumstances. Just as a heavy snow-
storm reveals a blockage in the coronary artery of the hapless shoveller, the
environmental crisis has revealed the unhappy state of ecology. The disease
of ecology is that it is a science that lacks theories it thought it had. When we
are asked to predict the impact of a new dam, or of an oil spill, or of chronic
chemical contamination, we cannot do it.
This diagnosis can be illustrated with examples from the greater literature,
but such critiques are available elsewhere (Peters 1991a, Schrader-Frechette
and McCoy 1993), so I need not repeat that information. This chapter dwells
on questions of lesser generality which arose in my own work. In part, I do
this because I know that work better, so I can criticize it more easily. I also
have a more subtle reason for self-criticism. I want to show that all ecologists
carry the infection and to encourage others to be at least as critical of them-
selves as they are of others.
Science and Ecology
Our problems arise because ecologists do not seem to appreciate fully
what a science should do. The job of science is to produce and examine a set
of one or more statements, called theories. A scientific theory is a tool that
specifies which of an infinite range of possible states are likely to occur, and
which are not (Fig. 9). In other words, a scientific theory is a statement that
makes a prediction and so tells us what we can expect to observe.
48 THE ECOLOGISTS’ DISEASE: TWO PERSONAL EXAMPLES

The most informative theories identify the likely future observations of the
system precisely and completely. To do this, they must exclude most
possibilities as unlikely. Thus the best theories are highly restrictive, and the
worst are slightly restrictive. Constructs that place no restrictions on the
future states of the system make no predictions at all and should not be called
theories.
We may want to do more than predict. For example, we may want to
explain or to understand nature. We may claim that we can do more, but those
claims only demonstrate our naivety. Science cannot do more. The only way
that we can demonstrate our understanding of nature is with predictive
power; all else is opinion and posture. By ignoring that basic limitation of
science, ecologists guarantee failure in their great endeavours, and direct
attention away from what science can do, prediction.
Non-theories
Some ecological constructs are called theories because they tell us what
might happen, even though they fail to identify what might not. As a result,
every observation is either consistent with the general construct or irrelevant
to it; no observation will falsify it. Such constructs are not theories at all. That
they are given this status by ecologists reflects our failure to think about
science.
The niche. The niche as a multi-dimensional volume in environmental
hyperspace (Hutchinson 1959) is a familiar example of non-theory. This
concept provides a model whereby we could order our observations about a
49NON-THEORIES
Fig. 9. A classification of putative scientific theories in terms of the degree to which
they eliminate possible future states of the system as improbable

species’ occurrence, or performance, as a function of some of an infinite
number of environmental factors. Because the hypervolume has no definite
shape, any form of the response will do. Because there is no minimum or
maximum number of factors to be specified, any set of appropriate observa-
tions can fit. Because there are always other axes to establish, the niche can
never be fully described, so seeming overlap between different species is not
unexpected. And because the significance of any degree of overlap on any
measured axis is unspecified, one is free to interpret overlap in any way one
wishes. In short, any observation is consistent with the construct, and none is
excluded. The multi-dimensional niche is wholly unrestrictive.
To assess a theory we must ask not “What facts are consistent with the
theory?”, but “What observations would be inconsistent with it?” That is
Popper’s criterion of falsifiability. If we focus where a theory may fail, we are
more likely to find its failings, to seek improvements and eventually to
develop better theories. But if we are interested only in showing how well a
theory performs, we will focus on confirming instances and entrench that the-
ory in the science, excluding alternatives and stultifying scientific advance.
We must remember that an infinite series of relations (i.e. of theories) can fit
any finite pool of data equally well (Chapter I), so confirmation is relatively
uninformative about the validity of a theory. As scientists, we must seek the
weaknesses of existing theory, so we can move to offer a better theory.
The competitive exclusion principle. The competitive exclusion princi-
ple provides a second case of an ecological non-theory. This principle goes
under several names, but most students of ecology are exposed to it in the
Lotka-Volterra equations. These equations ostensibly deal with the fate of
two competitors, Species A and Species B, occupying the same habitat. The
principle tells us that if you put two potentially competing species together
either A will be eliminated, presumably by competition, or B will be elimi-
nated, also by competition, or A and B will coexist, by sharing the habitat. A
final case, where both A and B die out, is implicit but uninteresting. The prin-
ciple is consistent with all possible outcomes; therefore it cannot be falsified
and provides no information.
Hardin (1961) recognized that the competitive exclusion principle was
unfalsifiable long ago, but he defended the principle as an aid to thought. For
example, it has caused generations of ecology students to puzzle through the
abstract algebra of the Lotka-Volterra equations, it has focused our thoughts
and research on examples of coexistence of similar species, and it allowed us
to organize observations of these examples within a common rationale, deter-
mining the resources over which competition is most intense, the likely
mechanisms of competitive exclusion, or those differences in resource use
that allow coexistence.
50 THE ECOLOGISTS’ DISEASE: TWO PERSONAL EXAMPLES

I pursued this example in my own work on the co-occurrence of calanoid
copepods in Ontario lakes. Calanoids are minute crustaceans that constitute a
major component of the zooplankton in most lakes, and in the sea. Hutchin-
son (1961) recognized that the co-occurrence of two or more species in the
homogeneous habitat presented by the open water of lakes represented a
major challenge to the exclusion principle. Co-occurrence of such similar
animals in such a simple environment should lead to exclusion, but it does
not. This “paradox of the plankton” became a classic example for limnologi-
cal applications of ecological ideas (Hutchinson 1951).
My first approach to the problem was to establish its reality. To do this,
samples were collected in over 100 lakes to determine the extent of coexis-
tence among calanoid copepods in Ontario. Calanoids were found in 95 lakes
and different species co-occurred in almost 3/4 of these. The animals were
represented by four genera: Senecella, Limnocalanus, Epischura and Diapto-
mus (Table 3).
We first used the principle to rationalize and simplify our observations.
The principle deals with competitive exclusion, but Senecella and Epischura
are thought to prey on other zooplankton; we therefore eliminated those gen-
era from consideration. As disciples of Darwin we had no difficulty in posit-
ing that ecological competition is a universal phenomenon or in accepting
Darwin’s statement that competition between closely related species is likely
more severe. We therefore further limited our analysis to the most common
and speciose genus Diaptomus, on the grounds that the others may be suffi-
ciently distinct phylogenetically that they no longer compete with Diapto-
mus. This had the further advantage of directing our attention away from Lim-
nocalanus whose little known food habits might include predation.
Diaptomus is considered a filter-feeding herbivore, although its food habits in
nature have actually been little studied. Subsequently, the genus has been
divided into several genera. I will not use this taxonomic nicety, but it could
be invoked to save the hypothesis.
51NON-THEORIES
Species Lakes Species Lakes
Diaptomus minutus 70 D. birgei 2
D. oregonensis 69 D. ashlandi 1
D. sicilis 9 Epischura lacustris 49
D. reighardi 4 Limnocalanus macrurus 11
D. sanguineus 2 Senecella calanoides 5
Table 3. The number of lakes in southern Ontario in which each species of calanoid was
found

After eliminating other genera from consideration, 92 lakes remain in the
sample; these contain seven common species of Diaptomus, and these con-
geners frequently co-occur (Table 4). D. oregonensis and D. minutus occur
together in about half the lakes, occasionally with a third species (D. sicilis in
six lakes, D. sanguineus in two, and D. ashlandi in one). D. birgei or D.
reighardi occasionally co-occur with D. minutus. This completed the first
phase of study. Directed by the competitive exclusion principle, potential co-
occurrence among congeners had been established and a scientific puzzle had
been generated.
The second part of the study was directed at the mechanisms by which this
co-occurrence was achieved, or equivalently, at the way species in the same
lake divide their habitats so that apparent co-occurrence is not real. I will only
give an impression of these results (Rigler and Langford 1967). Fig. 10 shows
that the most common putative competitors, Diaptomus oregonensis and D.
minutus, differ in their depths of maximum abundance, although there is con-
siderable overlap and in some lakes their vertical distribution is virtually
indistinguishable. When a third species is present, it consistently inhabits a
deeper stratum than either D. oregonensis or D. minutus, although it may
sometimes overlap with the two common species.
Our results are therefore consistent with the competitive exclusion princi-
ple: coexisting competitors do seem to divide their resources. However, we are
not yet justified in concluding that spatial separation is sufficient to allow co-
existence. The overlap between Diaptomus oregonensis and D. minutus seems
great, yet these species co-occur in half of our lakes. We should therefore
expect further differences between these species. Sandercock (1967) showed
52 THE ECOLOGISTS’ DISEASE: TWO PERSONAL EXAMPLES
Type of association
ABCDEFGH I J
D. minutus X XXXXXX
D. oregonensis XX XXX
D. sicilis XX
D. reighardi XX
D. sanguineus X
D. birgei XX
D. ashlandi X
No. of lakes 36 24 19 6213112
Table 4. Frequency of associations in which the various species of Diaptomus were
found to coexist

that the species are also temporally separated both in breeding and in diurnal
migration. These differences are more substantial, and again lead to the con-
clusion that the two species coexist because they have divided their resources.
In short, our investigation of competitive exclusion in Diaptomus ended
like all other investigations of the issue. Whenever two coexisting species are
studied, we find ecological differences between them, and thus we support
the competitive exclusion principle. When we find no difference, as when D.
oregonensis and D. minutus had similar vertical distributions, we cannot con-
clude that the principle was wrong. That observation, like the supposition that
both species use oxygen and phytoplankton, is irrelevant because the princi-
ple allows co-occurring species common use of many resources, provided
they divide at least one.
The problem is not yet finished. We have not yet shown that these differ-
ences are enough to prevent competition. We might make further observa-
tions to determine other differences in resource use, we might determine
limiting resources for Diaptomus, or we could manipulate the abundance of
53NON-THEORIES
Fig. 10. Vertical distributions of co-occurring species of the calanoid copepods (genera
Diaptomus, Senecella, Limnocalanus, Epischura) in several Ontario lakes. The length
of the horizontal bars at the base of each polygon gives the scale since each is propor-
tional to 5 copepods l
–1

one species to see how the other responds. The principle suggests much more
work we might do. It acts as a fruitful paradigm.
I do not believe that any further studies would disprove the principle of
competitive exclusion. In every case, the results would be confirmatory, or
irrelevant. This is a serious flaw. When we deal with a predictive theory, we
must be able to specify the observations that would be inconsistent with the
theory. In the case of competitive exclusion, this is not possible. Our study,
designed in light of the principle, could not have shown the competitive
exclusion principle to be false.
Ecologists seem to like constructs that fit all relevant observations. Like
Aristotle and his intelligible principles, we use observations to demonstrate
the applicability of our ideas, to shore them up against criticism, and to
extend them to new phenomena. As “theories”, these generalities are com-
pletely safe, and completely uninformative. Unlike Aristotle the scientist, we
do not use the general constructs to identify critical observations that might
cause us to change our general ideas. Indeed, we cannot do so because the
constructs are not susceptible to test. If we followed the simple rules of sci-
ence, non-predictive constructs like these would have been eliminated years
ago, and ecology would be less cluttered with nonsense.
Weak Theories
The best theories are those that tell us exactly what will happen by identi-
fying only one future state among many as probable. In doing so, they iden-
tify the many other future states as improbable. We can call these highly
desirable constructs “restrictive theories” (Fig. 9), because they restrict or
limit the range of possible futures to a narrow range of probability. Such pre-
cise predictions are not common in ecology, but they can be found where a
system is highly controlled or where the experimental manipulation is very
large.
Ecology contains many theories which are only slightly restrictive. They
might identify one or two future states that would be inconsistent with the
theory. Other weak theories are simply quantitatively inexact. They predict
trends: neonate size increases with maternal weight; species number de-
creases at higher latitudes; algae increase with nutrients; etc. These theories
are predictive, but their predictions are so uninformative that the theories are
virtually useless. They survive because ecologists do not demand better.
Strong science recognizes that falsifiability and predictive power are relative,
so each advance raises the standards for acceptable predictive power.
Evolution by natural selection. Other weak theories address only a lim-
ited range of phenomena, but are misleading because we act as though they
54 THE ECOLOGISTS’ DISEASE: TWO PERSONAL EXAMPLES

addressed many more. For example, the theory of evolution by natural selec-
tion may be only weakly predictive. I say “may be” because I want to be gen-
erous. The principle may not be a theory at all (Slobodkin 1968, Peters
1991a).
Natural selection is characteristically invoked to explain or rationalize the
variety of animals inhabiting the earth and their exquisite fit to their environ-
ments. In those contexts, there are four possible observations: maladaptation
and extinction, adaptation and survival, descent with modification, and an
evolutionarily stable state. These alternatives exhaust the possible future
states, but no general statement of the theory of evolution allows one to pre-
dict what will occur in a given case, or the average response in a set of cases.
Respecifications of the general theory may exclude some of these possibili-
ties and therefore seem to offer predictions, but close analysis of those cases
reveals only a narrower range of possibilities tautologically entailed by the
particularities of the case (Peters 1976).
The theoretical status of natural selection apparently depends on some
minor implications. Darwin thought that his theory would be shown false if
any organ were found which existed for the benefit of another species. Ruse
(1982) suggests that the theory of evolution predicts that some animals will
be difficult to classify and that palaeozoic mammalian fossils will not be
found. Whether or not these are predictions from the general theory is not
under debate here (see Peters 1991a). Instead I want to point out that the
absence of altruistic organs or the problems of taxonomists are not phenom-
ena that interest evolutionary biologists. If the generality of evolutionary
theory does not predict the phenomena of major interest, it is not theoretically
relevant to those interests.
A construct is not valuable just because it can be made to yield a predic-
tion. That only meets formal requirements of a scientific theory. Constructs
can be theories and still fail to tell us anything we want to know. Popper’s
demarcation is only a bare minimum.
Weak theories are misapplied when they are used in situations where they
eliminate no possibility. Like non-predictive constructs, concordances of
weak theories and data are used to organize or “explain” unpredictable ob-
servations in terms of the generality, and to extend the sway of these non-
predictive generalities. The frequency of such concordance is then used to
defend the general construct in areas where its predictive power is negligible.
The increasing popular technique of “hypothesis testing” in ecology (Fretwell
1975) often exploits this confusion to warrant weak theory, rather than to pro-
mote predictive power. The only protection against such abuse is to ask in
every instance where a theory is invoked, “What observation would have
falsified the theory?”
55WEAK THEORIES
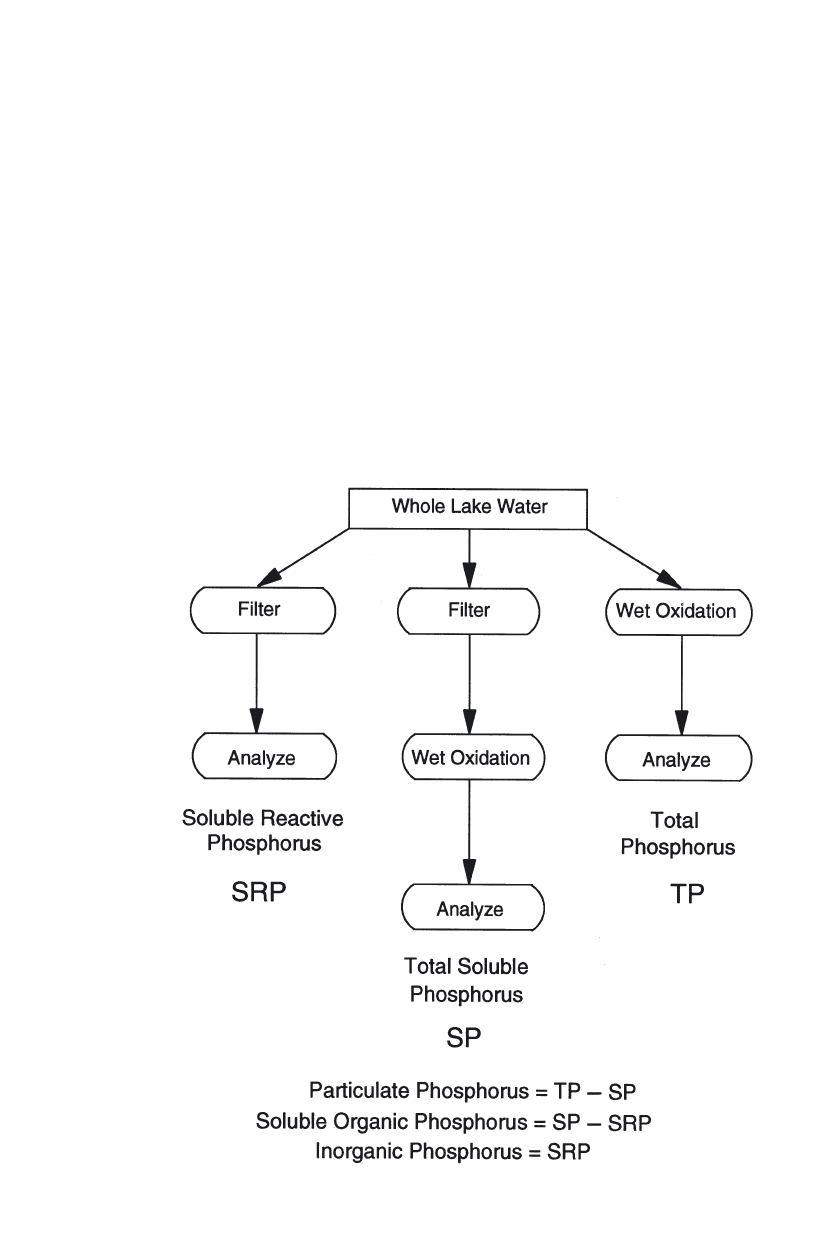
Concepts and measurement of phosphorus fractions. Because ecolo-
gists are uncertain about the nature of theory, they often seem to confuse
theory with fact. Established, or merely familiar, theories are treated as if they
were true, and therefore no longer worthy of the tests and scepticism that are
the due of every theory. Alternative theories are regarded as contrary to facts,
and falsifying facts are seen as erroneous theories. The effect is to entrench
error and to slow the pace of scientific response and growth.
I became aware of my own confusion of fact and theory in an analysis of
the various fractions that comprise phosphorus in lakes. Phosphorus fractions
are traditionally identified operationally (Fig. 11). To measure the “total
56 THE ECOLOGISTS’ DISEASE: TWO PERSONAL EXAMPLES
Fig. 11. The operations by which different fractions of phosphorus in lake water are
characterized

phosphorus concentration” in a water sample, the organic material in the sam-
ple is burnt away by wet oxidation. All the phosphorus in the sample is thus
converted to a soluble, inorganic form that reacts quantitatively with an acid
molybdate reagent to yield a blue compound. The concentration of this blue
compound is proportional to that of reactive phosphorus, so phosphorus
concentration can be measured spectrophotometrically (Griesbach and Peters
1991).
If the sample is filtered before analysis, the phosphorus in the filtrate is
taken to measure “dissolved” or “soluble” phosphorus in the lake water. Solu-
ble phosphorus is analyzed in two different ways. If the sample undergoes wet
oxidation before exposure to acid molybdate, the concentration of “total solu-
ble phosphorus” is estimated. If the sample is exposed to acid molybdate with-
out oxidation, the concentration of “soluble reactive phosphorus” is measured.
The difference between total soluble phosphorus and soluble reactive phos-
phorus is the amount of soluble phosphorus liberated by oxidation and may be
called “soluble unreactive phosphorus”. The difference between total phos-
phorus and total soluble phosphorus is a measure of “particulate phosphorus”.
Each of these fractions is subject to varying interpretations, but the most
common are as follows. Because ortho-phosphate (also called PO
4
or inor-
ganic phosphorus) also reacts with acid molybdate, without oxidation, solu-
ble reactive phosphorus is usually considered to be PO
4
; because ortho-
phosphate is a prime plant nutrient, soluble reactive phosphorus is usually
taken as a measure of available algal nutrient. Similarly, because soluble
unreactive phosphorus is not ortho-phosphate, it is often considered to repre-
sent biologically unavailable forms of dissolved organic phosphorus. Particu-
late phosphorus is thought to be a mixture of the phosphorus bound to living
and dead particles. Fresh water researchers now focus on total phosphorus
(Chapter IX), but many workers still see soluble reactive phosphorus as
particularly meaningful. Such considerations were apparent in the early work
at the Experimental Lakes Area in Ontario (Armstrong and Schindler 1971),
and still prevail in work by marine biologists and public health officers
(APHA 1989).
I became interested in these fractions when I began to wonder about the
biological availability of organically bound phosphorus, and sought a lake
with high concentrations of soluble unreactive phosphorus for my experi-
ments. The prevailing theory at the time was that soluble unreactive phos-
phorus concentrations were disproportionately high in tea-coloured lakes.
Hutchinson (1957) had advanced this hypothesis to explain differences in the
proportion of soluble organic phosphorus reported in lakes in Connecticut,
Michigan and Wisconsin (Table 5, overleaf). However, when I tested this
theory in Ontario lakes with different degrees of colour, I found similar
57WEAK THEORIES

proportions of soluble unreactive phosphorus in all lakes, despite changes in
colour (Table 6).
When I was unable to produce the expected colour effects, I looked for
alternative explanations, and noticed that previous studies had used different
techniques to isolate soluble and particulate phosphorus. Juday and Birge
(1931) had used a Foerst continuous centrifuge, Tucker (1957) had used three
layers of Whatman filter paper and Hutchinson (1957) “a 35-second mem-
brane filter”. I postulated that the reported differences in the amount of solu-
ble unreactive phosphorus might reflect these different methods, and tested
my hypothesis by applying different techniques to water from the same lakes.
I found that one could have almost any concentration of soluble organic phos-
58 THE ECOLOGISTS’ DISEASE: TWO PERSONAL EXAMPLES
Wisconsin Michigan Linsley Pond, Ontario*
Connecticut
(Juday and Birge 1931) (Tucker 1957) (Hutchinson 1957) (Rigler 1964)
Soluble inorganic 13 11 9.5 5.9
Soluble organic 61 48 28.5 28.7
Particulate 26 41 62.5 65.4
*Grenadier Pond was excluded
Table 5. The proportion (%) of total phosphorus in different soluble fractions and in
particulates as reported for lakes in Ontario and the literature. (From Rigler 1964)
Lake Total P % of total P in each form Colour
(µg l
–1
) (Pt units)
SRP SUP PP
Grenadier Pond 133 4.8 12.5 82.7 12
Heart 44 4.8 27.8 64.7 23
Teapot 33 5.0 29.6 65.2 42
Mary 27 6.8 25.0 68.2 11
Eos 18 4.8 28.1 67.1 162
Costello 12 7.2 28.8 64.0 30
Opeongo (South Arm) 7 5.5 31.7 62.8 20
Lake of Two Rivers 7 5.3 28.9 61.8 24
Found 5 7.8 30.0 62.2 6
Table 6. Water colour and the proportion of phosphorus in different fractions in Ontario
lakes. SRP: soluble reactive phosphorus; SUP: soluble unreactive phosphorus; PP:
particulate phosphorus. Colour is measured by comparison to the colour of platinum
solutions measured in mg l
–1
. (From Rigler 1964)

phorus one wished by using the appropriate technique (Table 7). Apparently,
the proportion of “particulate” or “soluble” phosphorus in a lake depends on
the technique used, presumably because a continuum of small particles and
colloids prevents any sharp distinction between the two fractions.
I had a very similar experience with soluble reactive phosphorus. Because
soluble reactive phosphorus reacts with acid molybdate as if it were PO
4
, it
was assumed to be the major form of nutrient phosphorus in lakes. When I
wanted to investigate the possibility that soluble organic phosphorus, mea-
sured as soluble unreactive phosphorus, was also a nutrient, I tried to strip the
PO
4
from filtered lake water with an ion exchange column. To determine the
efficiency of the column, I added a small amount of radioactive phosphate to
the water before passing the water through the exchange column. When I ana-
lyzed the elutriate, I had a surprise. Retention of soluble reactive phosphate
was high only when elutriate volumes were very small. When large volumes
of water passed through the column, more soluble reactive phosphorus
appeared in the elutriate. This did not seem to reflect exhaustion of the reten-
tion capacity of the column, because the efficiency of retention of
32
PO
4
remained high. Soluble reactive phosphorus behaved more like soluble unre-
active phosphorus than like PO
4
(Fig. 12, overleaf). This behaviour suggested
that soluble reactive phosphorus was not a measure of available inorganic
nutrient, and that the soluble phosphorus in lake water was almost all unavail-
able, despite its lability with acid molybdate, and the opinion of ecologists.
A further anomaly supported the view that soluble reactive phosphorus
was not available to algae. When a small amount of PO
4
is added to lake
water, it is rapidly taken up by the seston (Einsele 1941, Rigler 1956). Even-
tually, a steady state is established with 90 to 98% of the label in particles and
59WEAK THEORIES
Method of Heart Mary Thompson Average
separation 8 Jul 9 May 9 Dec
(38 µg l
–1
) (16.5 µg l
–1
) (18.2 µg l
–1
)
Foerst centrifuge 66 55 85 69
3-layer No. 44 Whatman filter 34 40 37 37
5.0 µm Millipore filter 32 42 52 42
1.2 µm Millipore filter 34 38 40 37
0.45 µm Millipore filter 21 39 40 33
0.22 µm Millipore filter 16 28 20 21
0.1 µm Millipore filter 15 26 13 18
Table 7. Effect of the method of separating seston from water on the percentage of total
phosphorus appearing as soluble organic phosphorus. Values in parentheses are total P
concentration in the lake. (From Rigler 1964)

the rest in solution. This process (Fig. 13) is most parsimoniously described
as a two-compartment exchange between a small soluble compartment repre-
senting PO
4
in solution and a large particulate compartment representing the
labile fraction of the total particulate phosphorus. This exchanging particulate
compartment seems to consist of bacteria (Currie 1990) and might be pre-
sumed to represent 10 to 20% of the total phosphorus. If
32
PO
4
is added to the
soluble compartment, then at equilibrium, the ratio of radioactive tracer in
particles to that in solution (
32
P
part
:
32
P
sol
) should be the same as the ratio
between the pools of exchanging particulate phosphorus and ortho-phosphate
(P
exchange
:PO
4
). This identity allows us to calculate the size of the exchanging
particulate pool from the equilibrium distribution of tracer and the concentra-
tion of soluble reactive phosphorus (SRP), as an estimate of ortho-phosphate
concentration:
P
exchange
= SRP × (
32
P
part
:
32
P
sol
) (5)
However, when these calculations are made, the concentration of exchange-
able particulate phosphorus is too large, sometimes several times larger than
the total phosphorus concentration in the water. One of the measured values
60 THE ECOLOGISTS’ DISEASE: TWO PERSONAL EXAMPLES
Fig. 12. Retention of soluble unreactive phosphorus (SUP), soluble reactive phosphorus
(SRP), and radioactively labelled inorganic phosphorus (
32
P-PO
4
) when different vol-
umes of filtered lake water are passed through an ion exchange column. SRP
behaves like SUP and not like PO
4

must be in error, and the most likely source of this error is that soluble reac-
tive phosphorus does not represent PO
4
.
Subsequent experiments showed that the accepted technique vastly over-
estimates biologically available phosphate in lake water (Rigler 1966) and a
number of authors have reached similar conclusions (Lean 1973, Peters 1977,
Tarapchak and Herche 1988, Bentzen and Taylor 1991, Taylor and Lean
1991, Karl and Tien 1992). It appears that in phosphorus-limited systems,
soluble reactive phosphorus is an irrelevant measure of nutrient phosphorus.
Nürnberg has shown that the two are similar only where concentrations are
high, above 10 to 30 mg m
–3
(Nürnberg and Peters 1984).
This re-evaluation of various phosphorus fractions is not simply a history
of methodological improvement. I see instead a typical programme of normal
science whereby concepts, like available free nutrient, soluble but unavailable
organically bound nutrient, and nutrient that is already biologically con-
sumed, emerged from a dominant paradigm to set puzzles for the scientist.
We wondered how we were to measure these concepts, and we assumed that
the quantities we measured had the properties of the concepts: reactive phos-
phorus is available to algae or phosphorus that passes a fine filter is in solu-
tion. Working within the paradigm, I showed some inadequacies of current
techniques, but did not question current concepts.
61WEAK THEORIES
Fig. 13. A two-compartment exchange model to describe the plotted data showing rapid
uptake of radioactive phosphate by seston in lake water, in the absence of any change
in ambient level of PO
4
as measured by SRP concentration

Although I was pleased with the tests as pretty bits of science, I now
recognize that the programme of research they represented was flawed. The
programme confused the things I measured (e.g. soluble reactive phosphorus)
with the concepts they represented (e.g. PO
4
and free nutrient) and the theo-
ries in which they were involved (e.g. algae can only use PO
4
), until my mind
slipped easily back and forth between fact, concept and theory, confounding
and confusing these very different entities. I was trying to find a measurement
that would correspond perfectly to a perceived truth, a paradigmatic concept,
without realizing that I had no way of recognizing this correspondence if I
found it. Apparently, I felt that the theory would arise almost automatically
from the data, once the concepts were perfectly measurable. I now believe
that those years of research, for all that they were interesting to me and pro-
ductive of publications, would have been better spent if I had instead asked
myself what theories could be built around the measurements I could make
and what predictions these theories would yield. I might then have discovered
much earlier that there was no theory waiting to use the concepts I was
studying.
62 THE ECOLOGISTS’ DISEASE: TWO PERSONAL EXAMPLES

V Broader Symptoms of The Ecologists’ Disease
“The environ is a device to truncate the infinite time regres-
sions and progressions propagating, respectively, inward
toward and outward from the holon center. The truncation
occurs at the border of the system to which the defining
holon belongs, and hence the environ is also relative to this
system. Accordingly, an environ is defined as a holon
together with its associated within-system environment.”
B. C. Patten
[Environs: Relativistic Elementary
Particles for Ecology (1982)]
Confusion of concept, theory, and fact has made any assessment of ecology
difficult, because the criteria on which scientific assessments should be
made are obscured. The resulting fuzziness makes research proposals hard to
judge, bewilders potential students, and obfuscates the few useful theories in
ecology.
This chapter continues the examination of ecology that began in Chap-
ter IV, and thus illustrates the disorder of ecology by pointing out some of the
symptoms. I begin by discussing our difficulty in posing large questions that
can be answered scientifically, because I believe that this difficulty has led to
our interest in relatively uninteresting minor problems that can be easily
addressed. I then examine the dissatisfaction, even contempt, that some ecol-
ogists seem to have for imperfect theories, for probabilistic statements and
simplifications. Next, I consider our tendency to let untestable concepts grow
unchecked and the characteristic lack of concern for scientific rigour in eval-
uating the theories we have. Finally, I consider the seeming incapacity of
ecologists to use quantitative theory effectively. In all these examples, the
underlying theme is that in ignoring the criterion of falsifiability, scientists
discard or fail to grasp the touchstone of scientific quality. This fault has
ramifications for the entire scientific enterprise.
Framing Scientific Proposals
I have generated two realistic, but artificial, introductions to research pro-
posals or scientific papers to illustrate how easily we let slip the opportunity

to pose relevant scientific questions (Table 8). These introductions might
have introduced the two personal research topics discussed at length in the
previous chapter. Both proposals begin by appealing to some large, generally
desirable goal, like feeding the hungry or understanding secondary produc-
tion. But both end by proposing a highly specific study whose relation to the
grand goal is obscure.
Given this mode of operation, whereby the projected work does not strive
to resolve the specified problem, almost any generally recognized societal or
scientific problem will suffice as background for a scientific project. Instead
of identifying a testable theory about the larger issue, typical ecological intro-
ductions point out that we need to understand, or that we are ignorant of,
some component. We cannot know if this component is really important,
because no theory specifies it as a critical element. Often there is only an allu-
sion to some shared belief and that allusion allows one to introduce a specific
topic (e.g. the relation between SRP and PO
4
, or the distribution of Diapto-
mus in Ontario) under the aegis of a grand one (e.g. feeding the hungry).
The jump from grand science to little science is hidden by reference to our
“ignorance” or our “need to understand”. Such phrases are like the black-out
of a theatre stage between scenes, for they allow a change of topic without
any logical inter-connection. If that intellectual sleight-of-hand is not obvi-
ous, it shows us to be enured to such tricks. We no longer find them surpris-
ing or disruptive. We no longer question what they might mean. If we are
actually ignorant, we cannot know enough to mount a scientific investigation.
If we seek understanding, we should specify how we will know when this
understanding has been achieved.
Once we accept the Popperian criterion of falsifiability, we know we are
not ignorant because we can specify one or more theories that restrict the
states that we are likely to observe. We would know when we have tested the
original hypotheses because we can compare observed and predicted states.
We would know when we have improved our original hypotheses, because
we could specify states considered likely under the old theories that are
unlikely in the new ones. And we would know that we have increased our
understanding when these new hypotheses survive further rigorous tests. In
the absence of hypotheses, introductions like those in Table 8 only indicate a
topic that interests the proponent.
The Reception of Moderately Restrictive Theories
Ecology has a number of empirical theories which are only moderately
restrictive. For example, given the average concentration of phosphorus in a
64 BROADER SYMPTOMS OF THE ECOLOGISTS’ DISEASE

65THE RECEPTION OF MODERATELY RESTRICTIVE THEORIES
Introduction
The world is hungry for animal protein. Most protein for human consumption is grown
on land, but this is inefficient, because farm animals compete with food crops for space
and because of inefficiencies in the conversion of fodders to homeothermic tissue. The
sea covers 70% of the planet but as yet produces only 10 kg of food per ha. Canada may
have five million lakes that are even less productive. Intensive aquaculture can yield
100 times more food. There is thus great potential, if the factors that limit productivity
in either fresh or salt waters can be relaxed. Of the two types of system, tractability and
small scale argue that it would be more efficient to first assess the determinants of sec-
ondary productivity in lakes, and then to use these results as models with which to
approach the limitation of productivity in the sea.
Proposal I
Nutrients usually limit the productivity of aquatic systems, yet the process by which a
given quantity of free nutrient is transformed into consumable flesh is virtually
unknown. Understanding this process will be the work of many scientists for many
years, but an important first step is the accurate assessment of available nutrient. In
lakes, phosphorus typically limits productivity and free phosphate is measured as solu-
ble reactive phosphate (SRP) by exposing filtered lake water to acid molybdate reagent.
A growing body of evidence suggests that SRP overestimates bioavailable phosphorus.
This project reassesses the relation of SRP as a measure of free phosphate.
Proposal II
Calanoid copepods form the base of pelagic marine food chains and are arguably the
most abundant organisms on earth. Yet they are only very imperfectly understood. In
large part, our ignorance reflects the difficulty in sampling and studying wild popula-
tions across thousands of square kilometres of open ocean.
Lakes can be studied more intensively and at less cost. Since calanoid copepods are
also important in lakes, the approximation involved in studying limnetic models of the
marine system is not extreme. We can therefore use lakes to develop and test initial
hypotheses about the structure and function of calanoid communities. Promising
hypotheses will eventually be tested in marine systems, but initial probing and sorting
of data and theories can be more efficiently followed in lakes than in the sea itself.
This programme will determine how congeneric species divide up a seemingly
homogeneous environment. We will survey a large number of lakes to establish the
degree to which species co-occur in different lakes and the amount of spatio-temporal
overlap in their distribution within lakes. If co-occurrence is as prevalent as anecdotal
and scattered literature accounts suggest, we will determine the likely factors that per-
mit these animals to share their simple habitats.
Table 8. Justifications for two programmes of normal scientific research in ecology.
After an ethically strong introduction, we can justify almost any study by invoking our
ignorance, the use of model systems, and the importance of understanding some funda-
mental part of the system. In neither proposal is it clear how the work will solve the
important problem in the common introduction

lake, it is possible to predict the mean chlorophyll concentration (Dillon and
Rigler 1974a) to within an order of magnitude, and given the body weight of
a Daphnia, one can make a similarly approximate estimate of its respiration
rate (Peters 1987). These moderately restrictive theories predict a range of
states, so the information they provide is probabilistic.
Probabilistic theories are good as far as they go, but we wish they went
much further. If we are planning a picnic, we want to be told that there is no
chance of rain, not that the chance is only 25%. Nevertheless, in the absence
of alternatives, such moderately restrictive theories have their place. They tell
us how well our science currently performs (as the explained variance), and
they indicate how much more we have to do (as the unexplained variance).
Moderately restrictive theories are not viewed as useful by all ecologists.
Some lament that the predictions are only approximate. For example, Shapiro
(1978) and Benndorf (1987) focus on the uncertainty of chlorophyll concen-
trations predicted from phosphorus concentration in lakes. They are con-
cerned because the chlorophyll concentration for a specific lake could lie
well off the value implied by the mean trend, and still be consistent with the
theory.
Others hold that such moderately restrictive, empirically based theories
are too simple. For example, the abundance of zooplankton, planktivorous
fish, piscivorous fish and spatial refuges can all be expected to affect chloro-
phyll development (Carpenter et al. 1985), yet they are not represented in pre-
dictive theories used for lake management (Chapter IX). The feeling that a
model is insufficiently complicated also reflects the confusion between what
contemporary science can offer and what we want. The critic posits the exis-
tence of a grand theory that describes the entire lake ecosystem and therefore
predicts chlorophyll effectively. This dream is compared to our present lim-
ited predictive power. If one discards the existing theory when it loses to a
phantom competitor, one is left with no theory at all.
These criticisms assume that explanatory theories spring fully formed
from the mind of their creator. That is not the lesson of experience. In de-
veloping their theories of astronomy and gravity, Copernicus, Newton and
Einstein drew on a tradition of recorded observation that dates back to the
beginning of history. The table of the elements was basically the description
of pattern in chemical behaviour; and the modern theory of the atom arose
from painstaking measurements of absorption spectra. Theory begins with a
search for pattern within the field of interest, passes to a rigorous description
or quantification of that pattern and, finally, to the incorporation of the pattern
into an explanatory theory. The idea that science can begin with comprehen-
sive explanatory theories betrays ignorance of how science has always
worked.
66 BROADER SYMPTOMS OF THE ECOLOGISTS’ DISEASE

Still other critics complain that moderately restrictive theories do not tell
us everything about the lake. For example, a theory that predicts mean sum-
mer chlorophyll tells us nothing about bio-accumulation of mercury (Lehman
1986a). Such complaints indicate a healthy desire for more and better theo-
ries. Such complaints are also disturbing because they indicate a general fail-
ure of ecologists to appreciate the nature of science and scientific theory. No
theory can do everything. A theory that predicts chlorophyll will not predict
contaminant burdens or algal taxonomy. A theory that predicts planetary
motion cannot predict the natural fall of a feather nor the flight path of the
bird that dropped it.
None of these counter-points to criticism should suggest that our current
empirically derived, moderately restrictive theories approach perfection.
They do not; replacements are needed and the current generation of theories
should be discarded as soon as better alternatives are available. The moder-
ately restrictive theories we have are only a step in the identification of pat-
tern (Fig. 14). But if we do not make some first step, we will never reach the
distant goals of better prediction and understanding for which so many of us
yearn.
The Pursuit of Ecological Concepts
Science progresses towards better theories by comparing the abilities of
competing theories to predict observations in critical experiments (Platt
1964). To make such comparisons, one must identify competing theories,
and devise experiments that provide sound tests. In ecology, critical tests of
competing theories are rare. Existing ecological theories are so little appre-
ciated that most ecologists would be unable to identify one theory to predict
67THE RECEPTION OF MODERATELY RESTRICTIVE THEORIES
Fig. 14. At present, empirical ecological theory is far from where we hope to go, but it
is a first step. (From Rigler 1982a)

a given phenomenon, much less two or more competitors. Unfortunately the
response to this deficiency has not been a search for ecological theories and
patterns.
Science uses theories to pinpoint observations of importance, and uses
observations to test theories. Ecologists rarely think in these terms. Instead,
we direct our thoughts towards abstract concepts that represent elements of
a sophisticated (but incomplete) mental model of the system we want to
study. If observations are used at all, they serve to confirm that the concepts
act in nature. Ecologists thereby abandon the practice that distinguishes sci-
ence and non-science, and our science suffers as a result. For example, that
approach identified free nutrient as a critical component in the phosphorus
cycle, and led me to two decades of work trying to measure a variable for
use in a future theory (Chapter 4) rather than testing a theory. The pursuit of
concept instead of theory is not just a personal failing. I share it with many
colleagues.
The limiting factor. The history of ecology is filled with abstract discus-
sions substituting for theoretical development. The concept of the limiting
factor is a case in point. In 1840, Justus Liebig proposed his famous “law of
the minimum”. He was concerned with the growth of plants and had observed
that a plant needs many different nutrients from the soil. If one of these nutri-
ents was absent, the plant would not grow. He formalized his observations in
the law of the minimum:
... if one is present in minimal supply, growth will be minimal. This is true
no matter how abundant the other foodstuffs may be. Growth is then
dependent on the amount of foodstuff present in minimal proportion.
At its inception, the limiting factor concept was applied only to the effect of
mineral nutrition on the growth of individual plants.
The next step was taken by F. F. Blackman in 1905 who was also con-
cerned with the factors controlling the metabolic responses of plants. How-
ever, he extended the concept beyond growth by considering other processes,
like respiration, and non-mineral factors, like temperature. Moreover, Black-
man described cases where high levels of the factor limited respiration, as
high temperatures reduced plant respiration. Thus, in Blackman’s work the
initial concept of the limiting factor began to expand and change. It got
muddled.
Shelford (1911) picked up Blackman’s concept and applied it to the distri-
bution of species: “The geographic range of a species is limited by the fluctu-
ation of a single factor (or factors) beyond the limit tolerated by that
species.” Shelford thus extrapolated the concept from a factor constraining
68 BROADER SYMPTOMS OF THE ECOLOGISTS’ DISEASE

individual metabolism to one determining species’ distributions. This exten-
sion may seem to offer wider application, but it also further inflates the con-
cept and renders any precise use of the term increasingly difficult. Elton
(1927) analyzed the geographic distribution of species more carefully than
Shelford and concluded that “biotic limiting factors” could act in conjunction
with abiotic limiting factors. Under that interpretation, a multiplicity of fac-
tors could act together, so the concept means little more than that the organ-
ism is affected by its environment.
F. E. J. Fry (1947) recognized the confusion gathering around the concept
of limiting factors. The concept seemed ready to treat all environmental
factors and all aspects of physiological and biogeographical response. To
address the problem, he reintroduced some rigorous terminology. For Fry, a
“limiting factor” was an environmental variable which regulates the
metabolic rate of an organism by virtue of its operation within the metabolic
chain. Thus nutrients, light, and oxygen could all be limiting factors, as
Liebig had suggested, but Blackman’s temperature, like pH, salinity, and
humidity, could not. Fry called these factors “controlling factors” which were
characterized by their ability to affect maximum and minimum rates, and all
rates in between. Finally, Fry recognized “lethal factors” that operated at the
extremes by killing the organism.
Fry’s redefinitions were rigorous and rational but they did not stem the
proliferation of conceptual meanings associated with the term. Eugene Odum
(1954) also began with a definition: “Any condition which approaches or
exceeds the limits of tolerance is said to be a limiting factor.” Thus Odum’s
limiting factors are equivalent to Fry’s lethal factors, and are nothing like
Liebig’s factors. However, when Odum uses the term limiting factor he obvi-
ously has many other definitions in mind. Sometimes his usage is consistent
with his definition: “Fire is ... an extremely important limiting factor”; but at
other points, his usage is similar to Fry’s: “In the geological and physiologi-
cal sense, the low concentration of CO
2
now existing is limiting to all land
plants.” Sometimes, he accepts that multiple limiting factors are at work, like
Elton, suggesting that “in lakes, oxygen, nitrate and phosphate are limiting.”
Later he introduces a new conception of limiting factors by claiming that “on
land ... wind exerts a limiting effect on the activities ... of organisms” and
illustrates the point with an experiment where the growth form of an alpine
plant was altered by a wind shield.
The limiting factor concept has continued to evolve. Hairston et al. (1960)
apply the term to an environmental influence that determines the maximum
population of a species. Limnologists refer to phosphorus as the factor that
limits primary productivity of a whole trophic level, or the secondary pro-
ductivity of a whole community. A glance at most contemporary ecology
69THE PURSUIT OF ECOLOGICAL CONCEPTS

texts reveals simultaneous use of multiple concepts. Each application of the
concept may be tied to observations, but these observations serve as confirm-
ing instances, not tests.
The end point of this development is that we can never hope to measure
“the limiting factor”. Ecological thinkers have associated so many meanings
and ideas with a single term that no measurable property could ever corre-
spond to all, and many different observations could correspond to one or
more ideas. Such a multi-facetted concept cannot play a role in theory, and
any construct that uses the concept, without rigorous redefinition and consis-
tent subsequent use, cannot be a theory.
This history of limiting factors is not an isolated case. Conceptual expan-
sion leading to terminological confusion and ending in unoperational concept
clusters is the common fate of many ecological ideas (Peters 1991a). As a
result, models that use such concepts cannot play a role in the formulation or
test of ecological theory. Ecologists who work with these concepts are exiled
to an untestable shadow-world that exists only in the imagination. The way to
avoid this limbo is to insist that the entities we discuss be measurable com-
ponents of predictive scientific theories.
Unconcern
Despite the developments outlined in the preceding section, some versions
of the limiting factor concept are among the most cherished simple theories in
ecology. Liebig’s law of the minimum says that the growth of an organism is
limited by only one substance at any one time. If more of that substance is
added, growth will increase, but if any other substance is added growth will
be unchanged. Extrapolation of the law to whole communities has had useful
consequences for eutrophication abatement programs: tertiary sewage treat-
ment and legislated changes to detergent composition have substantially
reduced phosphorus loads to lakes and rivers. The law of the minimum has a
strong intellectual appeal because it simplifies the theoretical problems of
complex systems.
Multiple limitation in the sea. In 1974, Theodore Smayda, a highly
respected oceanographer, published a paper in Limnology and Oceanogra-
phy, the foremost journal in his field. This paper challenged the law of the
minimum by concluding that in the nearshore waters of the western Atlantic
Ocean, as many as five factors could concurrently limit the growth of a single
clone of the planktonic alga Thalassiosira pseudonana.
Although Smayda’s paper claimed to falsify a theory that had been ac-
cepted for 150 years, it generated no excitement. It went almost as unnoticed
70 BROADER SYMPTOMS OF THE ECOLOGISTS’ DISEASE

as Gregor Mendel’s paper on inheritance. Each year, I look at the Science
Citation Index to see how many times the paper has been cited and why. And
each year it gains a modest few citations, a total of 40 by 1992, but no one
seems to appreciate that the paper challenges one of the dominant paradigms
in ecology.
Smayda’s paper is an example of the fate of heretical ideas raised at the
wrong time. Although Smayda’s credentials, choice of journal and style of
exposition followed accepted norms, Smayda’s analysis was received with
the same indifference as Velikovsky’s (Chapter III). This was to be expected
(Polanyi 1958, Kuhn 1962). Normal ecologists have quite enough puzzles to
keep themselves busy, and see no need to upset the world-view that justifies
those puzzles as scientifically interesting. Yet one would have hoped that
some supporters of the law of the minimum would have rallied to defend their
paradigm, if only because the prestige of the author and journal gave the con-
trary view some credibility. But no one cared.
There is an irony in this anecdote. If defenders of orthodoxy had attacked
the paper, they would have discovered a fatal flaw. I chanced to study the
article particularly carefully because I planned to use it in class as an
example of a sound bioassay. When I examined the data, I found only one
limiting factor, nitrogen. The paper compared growth rates in cultures where
all nutrients were in excess with those from cultures where all nutrients but
one were in excess, and found that the absence of any one of several nutri-
ents might reduce growth below the rate where all nutrients are present in
excess. Unfortunately, that finding is irrelevant to limitation in the field. The
proper comparison is with controls to which no nutrients were added.
Except for treatments where some toxicity was evident, growth rate in-
creased above that in the no-nutrient controls control whenever nitrogen was
added. If no nitrogen was added, growth rate was similar to those of the con-
trols despite the addition of other nutrients (Fig. 15, overleaf). Here is an-
other reason to suppose that the paper would have attracted serious atten-
tion, but again no one cared.
Most ecologists can point to other examples of flaws in experimental
design or interpretation. This example is therefore only one among many. I
do not intend to blame or discredit authors who have made mistakes in
their scientific careers. We all do that and should expect to do so. I am
instead concerned about a science which does not move to correct mistakes,
that offers no resistance to challenges to its principles, and that does not
read its own literature critically. Such nonchalance suggests a science deep
in normality and far from revolution. If we need a revolutionized ecology to
meet the environmental crisis, this evidence suggests that we are far from
ready.
71UNCONCERN

Inattention to Detail
Because ecologists are not used to working with theories, they often do
not realize that strong theories and predictions provide important checks on
poor measurements and miscalculations. If theory predicts one value for
some fact and observation yields a different value, there is an anomaly. Such
anomalies are fruitful. One might check the validity of the prediction, by
recalculation, and the observation, by repetition of the measurement. This
comparison allows us to identify human errors in both processes. If the
anomaly persists, one has a potential falsification of existing theory and the
basis for future scientific work.
In the absence of prediction, observational errors often go unnoticed and
the necessity for theoretical development is hidden. Under these conditions,
scientists lack a measure of validity, they may become sloppy and science
may stagnate.
72 BROADER SYMPTOMS OF THE ECOLOGISTS’ DISEASE
Fig. 15. Growth rate, expressed as a percentage of the maximum observed growth rate
of the alga Thalassiosira pseudonana (clone 13-1) exposed to sea water amended
with varying nutrient mixtures. Cultures which received all nutrients except nitrogen
(All – N) grew no better than control cultures (None), whereas cultures that received
nitrogen grew much better, regardless of deficiencies in other minerals. The variations
among cultures that received nitrogen may indicate other deficiencies that are ex-
pressed only when nitrogen is available in excess. TM = trace metals, VITS = vitamins.
(Data from Smayda 1974)

The calculation of secondary productivity. I became aware of this in-
attention in preparing a review of the methods of calculating secondary pro-
duction (Rigler and Downing 1984). This section is a precis of that review.
Secondary production has fascinated ecologists for much of this century
(Downing 1984). Many theories have been advanced to explain it and many
methods have been used to measure it. The basic concepts and techniques are
simple. Nevertheless, I found myself amazed at the total confusion that sur-
rounds these simple calculations. I worked through the literature in a futile
search for good examples of production studies. All I found was one error
after another.
I believe these errors arose because production biologists have been con-
fused by the many complicated and sometimes erroneous methods described
in the various instruction manuals and review papers to which they go for
guidance. I suggest that if we were to address ourselves to the simple princi-
ple underlying the calculation we would not make so many mistakes. Fur-
thermore, we would have more mental energy left over to devote to really
difficult and interesting problems.
The production of a population is the total amount of tissue that population
synthesizes during some interval of time. This production is a rate and has the
dimensions of mass or energy per unit of time. A year is often taken as the
unit of time, and for mass we might use fresh weight, dry weight, ash-free dry
weight, carbon content, nitrogen content, etc.
To illustrate the principle of calculating production, let us consider a single
cohort of an imaginary population (Fig. 16A, overleaf). It begins with a pop-
ulation of 10 individuals each weighing W
0
. After an interval of time t
1
, one of
these individuals, now weighing W
1
, dies. Its lifetime production is therefore
W
1
– W
0
. At a later time, t
2
, another animal weighing W
2
dies, and its lifetime
production may be calculated as W
2
– W
0
. This process will continue until the
last individual dies at t
10
and its production W
10
– W
0
is calculated. The total
production of the cohort over the interval from t
0
to t
10
is then the sum of the
final weights (∑W
i
) less the initial weight of the population (10 × W
0
):
P = (∑W
i
– 10 × W
0
)/(t
10
– t
0
) (6)
That is all there is to it.
Why then do we find whole books written on the subject of production,
and within them a whole series of different calculations? There are a number
of reasons that ecologists make such heavy going out of productivity calcula-
tions. One is that natural populations cannot be known as well as the imagi-
nary cohort in Fig. 16A. As a result, we must replace individual animals with
size classes, approximate the number of individuals in each size class and
73INATTENTION TO DETAIL

approximate the size limits of the class. The problems of approximation are
real and merit thought.
A second reason for our difficulties in treating secondary production is
that production ecologists erroneously believe there are three fundamentally
different methods of calculating production from these data:
(1) increment summation
(2) mortality summation
(3) the Allen curve.
These methods are essentially identical. They differ only in the way they sum
the weight increment over time (Rigler and Downing 1984). Each requires
74 BROADER SYMPTOMS OF THE ECOLOGISTS’ DISEASE
Fig. 16. (A) The exact calculation of secondary production by animals of known initial
and final weights. When the first animal dies, the production of the cohort is the area of
rectangle abcd; when the second animal dies, the cohort production is increased by the
area of rectangle defg, and so on until all the animals are dead, and the total area under
the curve is known. (B to D) Three variants in calculating secondary production of the
cohort in panel A. Dots indicate four observations of population size and individual
weight taken over the course of the cohort’s existence and represent the means in a size
class or over a time period. Heavy lines indicate the areas which are used to calculate
total production and shaded areas are errors in estimation
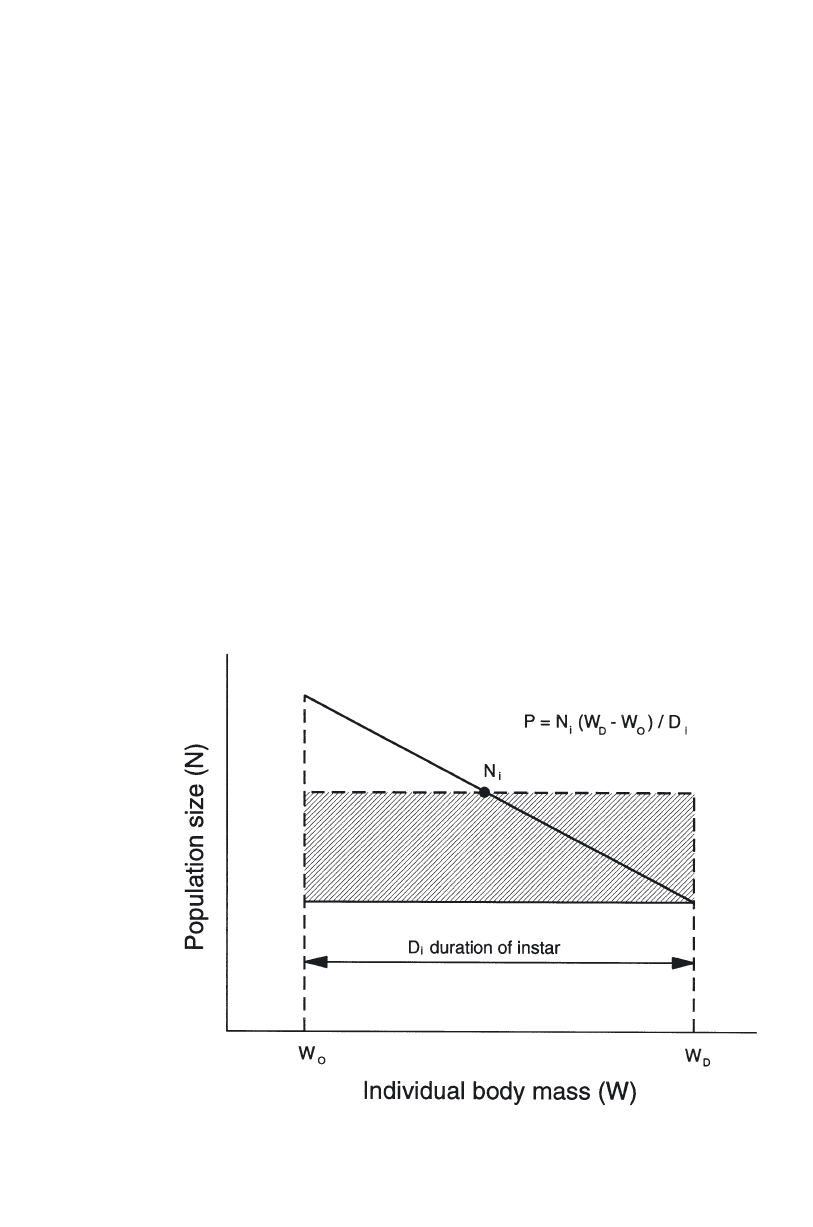
that the population be divided into a number of size classes, and that for each
size class one know N
i
(the average number of animals entering size class i),
W
min.i
(the mass of the average animal entering the size class), and W
max.i
(the
mass of the average animal leaving that size class; this is also the mass of the
average animal entering the next size class). Production is then calculated as
P = ∑N
i
(W
max.i
– W
min.i
) (7)
At this point, the interesting problems begin. Most real populations do not
produce cohorts. When we measure the number of animals in a size class over
time, we do not see a regular decline in numbers (Fig. 17), but a more or less
erratic pattern of peaks and troughs as animals are born, grow and die. If the
animals can be aged, they can be sorted by age class and treated as if they rep-
resented a cohort. If this is not possible, then production can be estimated
from the number of animals in the size class (N
i
), the upper and lower size
limits of the class (W
min.i
, W
max.i
) and the average duration of that size class
(D
i
):
P = ∑N
i
(W
max.i
– W
min.i
)/D
i
(8)
75INATTENTION TO DETAIL
Fig. 17. Approximation of the production of a cohort, age- or size-class, or population
based on a single estimate of N, estimates of minimum and maximum size (W
min
and
W
max
), and estimated duration of the size class (D). The magnitude of the errors in this
calculation depends on the schedules of mortality and growth over the period D, as well
as on how representative the estimates of D and population size are

This equation is really an extension of the model in Fig. 16 pushed to the
bare minimum of sampling effort (Fig. 17). It is an approximation to the
extent that Eq. (8) departs from Eq. (6). For example, Eq. (6) assumes that the
timing of growth and mortality are known for every animal, but Eq. (8)
assumes that they can be approximated by averages. In fact, changes in the
scheduling of mortality and growth over the interval D
i
could cause signifi-
cant errors in estimating production. For example, the production of animals
that died before sampling to estimate N
i
is left out of Eq. (8), and if these are
many, then the equation underestimates production. Conversely, the equation
assumes that animals alive at sampling survive to reach maximum size, and to
the extent that they die soon after sampling, the equation will overestimate
production. Finally, Eq. (8) assumes a linear growth curve, but if growth is
exponential, most of the production will occur towards the end of D
i
, when
population is smallest; and again Eq. (8) will overestimate.
For fish or clams or humans or trees, D
i
is easily measured, but for the host
of tiny invertebrates that dominate aquatic ecosystems, this determination is
not easy. Consequently, a number of so-called methods of calculating pro-
duction are really disguised attempts to avoid measuring D
i
. These attempts
almost always lead to error and add to our confusion. I will give only one
example: the size-frequency or Hynes method.
The Hynes method really excited stream ecologists because they could
never sample their animals well enough to calculate D
i
and, even when they
had their samples, they could not identify the species. They were accordingly
very pleased when offered a method that required neither D
i
nor identification
of species. This method is based on two implicit assumptions: that total devel-
opment time equals one year for all species and that the development time of
each size class or instar, D
i
, is a constant. That is to say, the method assumes
that animals spend the same amount of time in each size class. The
Hynes method therefore does not overcome difficulties, it merely ignores
them. But you cannot avoid measuring N
i
, D
i
, and (W
max.i
– W
min.i
). If you try,
you will go wrong and confuse everyone else. More problems are discussed
in Rigler and Downing (1984).
Let me summarize this section. Production has been considered important
by a generation of ecologists. The method of calculating production is so
incredibly simple that a child in elementary school could be taught it. Yet
almost every zoologist who has calculated secondary production has done it
incorrectly. There are similar errors in calculating primary production, and
probably in most quantitative treatments in ecology. We can take a lesson
from this example and that lesson contains both good news and bad.
First the good news. Ecologists are still frightened by the most elementary
mathematics. Consequently we use the literature like a cook book: we look up
76 BROADER SYMPTOMS OF THE ECOLOGISTS’ DISEASE

a method and use it without understanding the principle behind it. I know
personally that when I have to look at an equation and try to understand it, I
usually fail. I give up, feeling frustrated and stupid. The lesson in this section
is that if one has really tried to understand a paper and failed, the reason for
failure may not be that one is stupid, but that the paper is wrong.
Now for the bad news. I think the persistence and frequency of error in the
secondary production literature and in the rest of ecology shows that, by and
large, ecologists do their research superficially and carelessly. We act as if we
do not care about the validity of our results. We do not care because we are
not testing theory, so that any value will do. This is a sign of the sickness in
ecology.
We should all take pains to remove error and nonchalance in our own
work. However, we should also be concerned with the cause of these symp-
toms. If we could understand their source, we would make an enormously
greater contribution to our discipline. For me, the root of our problems is our
disinterest in prediction. Predictive theories require accurate measurements
to provide adequate tests and to avoid spurious anomalies.
Some Consequences
This chapter suggests that much of ecology is confused in its goals, uncer-
tain of its strengths, and inconsistent in its terminology. It portrays ecologists
as nonchalant about their tests, careless in their measurements, yet closed-
minded in considering alternatives. It claims that ecologists do not recognize
the few good theories they have, preferring to pass their time with weak theo-
ries and non-theories that substitute a form of explanation for prediction. It
also claims that ecologists rarely try to make predictions, believing that some
special property of their systems makes the common model for building and
testing theories inoperable.
This is an unattractive picture. It is one in which few ecologists see them-
selves. Yet I believe it is the face we turn to the public, to the granting agen-
cies, and to our students — with devastating results. In each case, a poten-
tially friendly and supportive audience is baffled by arcane subject matter.
Many turn away, feeling bored, frustrated, stupid, embarrassed and incapable.
I submit that the fault is not with them; they have sensed the real nature of
much of contemporary ecology.
Some students have such interest in ecology that they refuse to drop out.
Those few survive to become professors and researchers. Unfortunately, they
often do so by making the illogical jump from grand concerns to picayune
experiments. When they do so, they become part of normal science. They
77SOME CONSEQUENCES

accept the dominant paradigms of ecology and no longer see its anomalies
and failures. They become us.
This process of acculturation explains the failure of ecologists to appreci-
ate the nature of their science. We have all been brain-washed. To escape, we
must step outside the comfortable limits of normal science to see what needs
to be done, and to look back at what we are doing. We have to contemplate
our science.
78 BROADER SYMPTOMS OF THE ECOLOGISTS’ DISEASE

VI Why Limnology?
“There is nothing — absolutely nothing — half so
worthwhile as simply messing about in boats.”
K. Grahame
[The Wind in the Willows (1966)]
Science has a way of overwhelming its participants. Fields of interest that fit
within our scope during periods of youthful exuberance double in size every
ten to fifteen years (Price 1986), and even rare demonstrations of personal
competence invite additional duties. To succeed in their research, most con-
temporary scientists therefore have to dedicate themselves to the tasks at
hand: experimental design, observation, data analysis, publication and read-
ing in their specialty. One result is that scientists have little time to address
philosophical generalities or to worry about how their efforts contribute to the
larger body of science.
Our behaviour is justified if we believe that science is sufficiently self-
correcting: a proper overall direction arises automatically from the process of
thousands of researchers “doing science”. A similarly blissful ignorance fos-
ters the view that because any piece of scientific information is intrinsically
valuable, there is no need to fit the piece we study into a larger picture. We
may recognize the value of the long view, but too often our response is sim-
ple regret that we have not the time to think about it. None of this is accept-
able. We must reject the notion that scientific forethought is an unnecessary
luxury. To do otherwise is to deny the most fundamental of scientific tenets:
that humanity can achieve some power over our destiny by thought and
observation.
In this essay, I contemplate my own branch of science by trying to answer
a question that has bothered me for some time: Why teach or study limnol-
ogy? My answer is still only preliminary. I hope that by asking it, I will
encourage others to complete the process of analysis for limnology and to
extend that analysis to other branches of ecology, biology, and science.
What is Limnology?
Limnology is the scientific study of fresh waters. Within that limitation, its
subject matter is broad. Limnologists may be physicists, chemists, mathe-

maticians, geologists, hydrologists, engineers, or any of a host of other
specialists. Limnology could involve studies of the evaporation rate of water
collected by a pitcher plant, the thermal stratification of Lake Michigan, the
chemistry of iron, etc. For biologists, like me, limnology is more restricted:
limnology is the study of living organisms that inhabit fresh water. I will
restrict it further and deal only with living organisms in lakes. The reason for
this second restriction is partly personal, and partly convenient. I work with
lakes and like them, and if I demonstrate that studying the biota of lakes has
value then I will have demonstrated that limnology as a whole has at least that
same value.
There are obvious practical reasons to study fresh water. Water is one of
our most important and valuable resources. In Canada, many of us live in one
of the most famous lake districts in the world: five of the world’s largest lakes
lie at our doorsteps, and millions of smaller lakes dominate the landscape of
the Canadian shield and condition our lives. In the country as a whole, gov-
ernment figures suggest that fresh waters add 10 to 20 billion dollars to the
economy every year (Science Council of Canada 1988). In the United States,
extensive irrigation has allowed the deserts to bloom and feed a hungry
world. In England, a tenth of the waters of the Thames passes down domestic
drains on its way to the sea and at least as much again is used industrially. In
the developing world, water-borne diseases are major killers. At a practical
level, the need for limnology is obvious: fresh water is critical to industry,
agriculture, health, and nature, yet it is in short supply and its quality is
increasingly compromised. We must learn how to improve, to conserve and to
use this precious resource (Likens 1992). That is the practical challenge for
limnology.
Practical considerations are critically important, but they are not of central
concern in this chapter. Practical issues are not used in universities to justify
hiring a limnologist or offering a course in limnology. As university students
and teachers, we should be primarily interested in science, and the reasons for
hiring and teaching should hinge on the scientific value of the discipline. As
students, we should be asking “What is science?” As teachers, we should be
trying to answer that question. And as scientists, we should all be trying to
make a contribution to the development of science.
What is Science?
Before we can ask “Why limnology?”, we have to ask “What is this thing
called science?” Since I have discussed the point in earlier chapters, I can
answer arbitrarily here. Science is the process of making, testing and using
80 WHY LIMNOLOGY?

theories. These theories are verbal or mathematical statements that make pre-
dictions, and thereby give a functional description of the world in which we
live. I say “functional” because our theories need not give a “true” description
of the world. In fact, philosophers have shown very convincingly that science
cannot hope to give a true picture of the universe.
Our job as scientists is to produce a set of statements or theories that are
consistent with each other and which make good predictions about future
events and observations. In other words, the essence of science is in its theo-
ries, and the justification for any and every piece of research lies in its rela-
tion to a particular theory. If someone asks what you do, it is useless to
answer, “I am an entomologist”, or “I am a limnologist”, as this describes
only the tools of research. Insects and lakes are only the material used to test
theories. A better answer would be “I am interested in gene theory” or “I
study competition theory”, because this describes the ideas you wrestle with
in your research. I define ecology as the science that predicts the abundance,
distribution and other characteristics of organisms in nature. Biological lim-
nologists, as freshwater ecologists, seek to predict these characteristics of
organisms inhabiting fresh water. If limnology is to interest other ecologists,
it must show them how the study of lakes relates to the tests and problems of
other fields.
Now we can return to our original problem: “How do we justify limnol-
ogy?” We now know that we can justify it only by showing how limnology
allows us to test a scientific theory or a group of theories. In effect, we now
know what to look for among all possible justifications and our search will be
much shorter. We are looking for theories related to the study of organisms in
their natural environment, and I will argue that lakes are a particularly good
place to test those theories.
Ecological Theories
There are many theories waiting to be tested in ecology. We have theories
about how individuals use their resources, about how predators interact with
their prey, about how populations compete, about how food webs are orga-
nized, about how higher trophic interactions affect plants and about how
variations in plant nutrients affect predators. In short, we have many, many
theories about how organisms interact with their environment.
Where will we test these theories? What constitutes the environment?
How extensive is the population? or the community? Do we consider the
assemblage of species in the whole world as “living together”? Are there
practical limits to the system we wish to study? Perhaps it is hard to imagine
81ECOLOGICAL THEORIES

that an Australian earthworm and a North American beaver share any res-
ources, but does this mean we should treat continents as our units? If not, how
do we carve the world up into ecological units? Are we actually justified in
dividing it up at all? These questions have always been thorny ones for ecol-
ogists and they show no sign of resolution. Modern chaos theory has given the
problem new respectability as “the butterfly effect” whereby small causes
may have grand effects, as the reverberations from a butterfly’s wing in
Africa may determine weather patterns half a world away (Gleick 1987).
The ecosystem concept. Ecologists believe that we can and, in fact, must
divide the world up in order to study it. We call this “the ecosystem concept”.
The idea that we could identify lakes as discrete ecological units was formu-
lated back in 1887 by Steven Forbes in The Lake as a Microcosm. This article
had such a strong influence on amateur as well as professional biologists, that
it was reprinted at least twice and is still cited in the literature. Forbes begins
his paper:
A lake is to the naturalist a chapter out of the history of a primeval time,
for the conditions of life there are primitive, the forms of life are, as a
whole, relatively low and ancient and the system of organic interactions by
which they influence and control each other has remained substantially
unchanged from a remote geological period.
The animals of such a body of water are, as a whole, remarkably iso-
lated — closely related among themselves in all their interests but so far
independent of the land about them that if every terrestrial animal were
suddenly annihilated it would doubtless be long before the inhabitants of
the lake would feel the effects of this event in any significant way. ...It
forms a little world within itself — a microcosm within which all the ele-
mental forces are at work and the play of life goes on in full, but on so
small a scale as to bring it easily within the mental grasp.
Forbes makes a number of interesting points in this short section of his
paper, but I want to draw attention to only two: organisms in a lake are un-
affected by those on the land; and the ecological processes in a lake are so
primitively simple and the scale of these processes is so small that the lake
system can be easily comprehended.
These two ideas have been very influential. They have attracted many
ecologists to work on lakes, and convinced many limnologists that the study
of lakes is more likely to be rewarding than the study of streams. If the organ-
isms in the lake are isolated from events and organisms on the surrounding
land, then characteristics of the lake alone must determine the properties of
the lake system. If lakes are simple systems, they will allow us to see general
ecological principles in action. Thus a lake provides ecologists with a simple
model system in which to test their ideas.
82 WHY LIMNOLOGY?

Forbes and his followers saw that it was possible to divide up the bio-
sphere into little, self-contained units in which all ecological processes could
be observed. He called this unit a microcosm, but many terms have been used
for this concept. Nowadays, everyone uses the term ‘ecosystem’. English-
speaking writers usually attribute the term to Tansley (1935), but Remmert
(1980) mentions that it was introduced by R. Woltereck a decade earlier and
the concept is older still.
An ecosystem is an ecological unit comprising living and non-living com-
ponents interacting to produce a stable system. An ecosystem might be a
forest, an area of grassland, a stretch of desert, an oceanic island, a temporary
pond, an alpine meadow, a lake, or any other piece of the biosphere. The
ecosystem has become a basic unit for ecologists, just as the atom is a basic
unit for chemists and the species is a basic unit of taxonomists. Since the
ecosystem is considered to be the smallest unit in which all ecological pro-
cesses occur, it may be the functional unit of the biosphere (Odum 1971). If
so, ecologists must eventually test their theories in ecosystems, and until a
theory has been shown to be relevant in a real ecosystem, its relevance to
ecology is speculative.
Why limnology? Now we can return to our original question: Why study
limnology? And since I have explained why we should study the ecosystem,
our original question can be resolved to
“Why study this particular ecosystem?” Why not offer a course in grass-
land biology, forest biology or even urban biology? The answer, if there is
one, should be that the lake has properties that make it a more suitable eco-
logical tool than other ecosystems. At least at first sight, lakes offer many
advantages for testing theory, and I believe that these advantages explain
much of the early interest in limnology by ecologists. So let us now look at
some characteristics of a lake that make it a better model ecosystem than
many others.
A terrestrial ecosystem. Instead of looking at lakes in isolation, let us first
consider a wood bordered by grassland (Fig. 18, overleaf) to see the limita-
tions of terrestrial ecosystems as objects of study. The wood seems a self-
contained ecosystem; it includes a whole assemblage of plant species not
found in grassland, and many mammals and birds found there do not enter
surrounding grassland. Within this woodland, nutrients are cycled, energy is
fixed and degraded, predators prey and plants reproduce. It is an ecosystem.
But let us look more closely. Ask how the oak interacts with the beech. Do
they share nutrients or energy? Would removal of beech affect the oak in any
way? One can always argue that there might be an effect. Perhaps some squir-
rel that depends on nuts from both trees is hiding them together so their off-
spring compete. My point is not whether there are or are not interactions, but
83ECOLOGICAL THEORIES

only that some of these interactions appear weak and tenuous. It is easy to
imagine that interactions within the woodlot are no stronger than those
between the beech tree and the nearby shrubs or grasses at the edge of the
adjacent grassland ecosystem.
Now consider large herbivores or predators associated with our ecosys-
tem. How do we handle the fox that hunts mice and insects in the wood, the
grassland and the adjacent woods across the valley? What do we do about the
male robins that come from miles around to spend each night in a bachelor
tree, leaving behind little heaps of concentrated nutrient? We seem to be run-
ning into problems with our terrestrial ecosystem. Its boundaries have be-
come so blurred that we begin to wonder if it is really there at all.
Finally, consider the practical problem of sampling a terrestrial ecosystem.
Except in the very special cases where the researcher can count and measure
each individual, every ecological study begins by sampling the populations of
some species. In a forest, this is not easy, for each group requires special tech-
niques. Plants are not evenly distributed and must be sampled in randomly
selected quadrats, deer may be sampled by aerial surveys, mice by capture-
recapture with live traps, soil invertebrates by Berlese or Tullgren funnels,
moths by night-lighting, and other insects by sweep-sampling or ground-
sheets. Estimates for mobile species are often affected by emigration and
immigration, and some measures of abundance are only indices that cannot
be compared to each other.
This hypothetical example and its suite of rhetorical questions highlight
the problems of applying the ecosystem concept to real ecosystems. Interac-
tions, such as the sharing of energy and nutrient, between any two organisms
within same ecosystem are sometimes less than those between two organisms
84 WHY LIMNOLOGY?
Fig. 18. A woodland ecosystem

in adjacent systems. Animals migrate from one system to another and we
must decide where they belong. The transition from forest to grass may be so
gradual that it is difficult to say where one ecosystem begins and the other
ends. An added difficulty is that the assemblage of species in the oak-
dominated hilltop differs from that in the beech-maple forest lower on the
slopes; one is always tempted to divide the single ecosystem further.
A limnetic ecosystem. The confused and complex situation of the terres-
trial ecosystem contrasts sharply with the apparent simplicity of a lake
(Fig. 19). The first difference is that the lake has definite boundary: a step in
one direction puts you in, and a step back gets you out. Some animals, like
kingfishers and frogs, cross the boundary, but because conditions change so
abruptly at the water’s edge, most do not. Boundary-crossers are much less
common than on land.
The second difficulty we encountered with a terrestrial ecosystem was that
it is sometimes more difficult to show interactions between individuals within
one ecosystem than between individuals in adjacent systems. Now look again
at lakes and see if this difficulty exists (Fig. 20, overleaf). The lighted, well-
mixed, surface layer of a lake is called the epilimnion. It is here and in the
upper portion of the next layer (the metalimnion) where most photosynthesis
takes place. The photosynthesizers in this ecosystem are not trees, but minute
algal cells, 1 to 200 µm in length. Now consider cells at two positions (A and
B) in the epilimnion, and ask if we can expect interaction. Because the wind
continuously stirs the epilimnion, the algal cells there can be compared to
lumps of vegetable in a stew. Each leaks organic material into the medium
and each absorbs the flavour of the others. If you stir long enough each lump
will bump into every other lump. We can therefore conclude that interactions
are potentially very strong and that all cells compete for nutrients and energy.
A related advantage to this stirring is that it imposes considerable horizontal
uniformity on the system. Just as we have less trouble distinguishing bound-
aries, we are also less tempted to divide the system further.
The third difficulty concerned sampling terrestrial systems. Now consider
lakes. The plants are microscopic, many cannot swim against the current and
85ECOLOGICAL THEORIES
Fig. 19. A lake ecosystem

their horizontal distribution will usually be sufficiently uniform that a single
vertical series of samples will give a good estimate of biomass (Hanna and
Peters 1991). Zooplankton are more mobile and may be affected by wind so
vertical samples must be taken at different positions in the lake (Langford and
Jermolajev 1965, Prepas and Rigler 1982), but this is still a simple procedure.
Sampling fish is a more onerous exercise, but because migration is negligible,
the method of capture-recapture is probably valid.
There are other advantages to lakes as systems of study. Aquatic commu-
nities are dominated by minute organisms with short lifespans; such organ-
isms can respond almost instantaneously (at least to the human observer) to
disturbance, so the influences of an unknown history are presumably much
less than in terrestrial communities, and the likelihood of encountering a
quasi-steady state seems larger. If we avoid the discontinuities associated
with the surface and the mud, the lake community is more homogeneous than
the forest or grassland, so that sampling can be less intensive. Horizontal
changes are small and existing vertical gradients in light, temperature and
oxygen are gentle and predictable.
I will stop here because my object is not to convert all ecologists to lim-
nologists. I am not proselytising for limnology, but for a way of approaching
research. A scientist’s job is not to make measurements that no one has made
before, it is to construct and test theories. Science always begins with theory,
whether with a theory that needs testing or with groups of facts that should be
related by a theory. You should try to discover where your interests fit into the
scheme of things in science by showing how your pet theory is related to
other theories. When you test predictions of your theory, look for the system
— whether a cell, a species or an ecosystem — that makes your job simpler.
A difficulty arises because aesthetics almost always enter into biological
research. We are biologists because animals or plants happen to please us.
One researcher likes fish and finds them beautiful, but hates birds; another is
86 WHY LIMNOLOGY?
Fig. 20. Gradients in light and temperature in the lake ecosystem. A and B are two
distant points in the epilimnion

just the opposite. Personally, I find lakes and the organisms they contain
aesthetically pleasing. Obviously, one should not try to test a theory with
inappropriate material, however fascinating you think it to be; but equally
obviously, we will not give up the subject we like to work on for one we find
repulsive, just to test a theory. If we begin with the subject matter, we must
ask what biological theory or theories can best be tested with that pet mate-
rial. Thus we still look for the theory before we start making measurements.
Ecological theories relate an object of particular interest to its environ-
ment. In lakes, both the object and its environment are easily defined. This
practical detail, coupled with the theory that the lake ecosystem is easier to
understand, made the lake ecosystem an important tool in the search for and
the testing of ecological theory.
A paradigm shift in limnology. This argument for limnology explains the
interest of early ecologists in limnology, but not its current role. Some of us
still see limnology as a vital field, providing leadership to the rest of ecology.
However, for many scientists both inside and outside limnology (Jumars
1990, Wetzel 1991), limnology has lost its central position in ecology. There-
fore I want to end this eulogy to my beloved science with a further sugges-
tion.
The early lead that Forbes gave to limnologists allowed us to discover the
limits of the ecosystem concept before other ecologists. The idea of the lake
as a microcosm now threatens to be restrictive rather than instructive. Lim-
nologists who have gone beyond that paradigm are now poised to enter a new,
more powerful phase of scientific development in which limnology will once
again be a major branch of ecology.
The lake is not a microcosm. In Europe, limnologists were less influenced
by the beauty of Forbes’ English (Elster 1958). Some held similar views
(Pearsall 1932), but others disagreed. Thienemann (1926) stressed the role of
lake morphometry in determining the composition and abundance of organ-
isms in lake communities. A deep lake would be unproductive and rich in
oxygen; shallow lakes were instead productive and their deeper waters were
deficient in oxygen. Both types of systems would have the flora and fauna
appropriate to their respective conditions. Naumann (1930) instead argued
that lake properties were determined by the geology of their basins.
At first, limnologists on both sides of the Atlantic were little concerned
with the impact of the land on lakes or with the evolution of lakes under these
impacts. Because the life of lakes is long relative to our own, we first thought
of lakes as we first thought of species, as permanent. Then paleobotanists
began coring bogs to reconstruct the history of terrestrial vegetation, and
found that bog sediments contained the remains of open-water algae. This
discovery forced us to extend our time scale and to admit that lakes change
87ECOLOGICAL THEORIES

and die. We began to think of the evolution of lakes as similar to the life of a
star: A lake is created in a catastrophic event such as the advance and retreat
of a glacier; it develops strength as it slowly accumulates nutrients; it ages as
sediments accumulate; it gradually fills, becoming shallower and richer; and
it finally dies in a eutrophic burst of productivity before it is transformed into
a meadow of sedges and grasses. This view of lake evolution was basically
consistent with the ideas of Thienemann.
As the human population increased, human observers first on the shores of
Swiss lakes (Ambühl 1960, Thomas 1969) and then on the shores of North
American lakes (Beeton 1969, Edmondson 1969) began to see changes within
their own lifespans. We had to admit that the evolution of a lake was not so
slow as we once thought. In fact, only a few lakes, like Baikal, Titicaca, and
the African Rift Valley lakes, antedate the pleistocene. The rest are only
10000 to 100000 years old. The inhabitants of these lakes cannot be the rem-
nant from a distant past. Moreover, we had to admit that climate and human-
ity could hasten the aging process and affect the properties of the evolving
lake. We began to realize that lakes were not isolated systems. They were very
much influenced by what happened on land. We became interested in pre-
dicting these processes, and much of the current support for limnology is
directed to predicting the effects of development in the drainage basin on the
properties of the lake.
Evidence against the hypothesis that lakes are isolated ecosystems is now
everywhere. The watershed and airshed bring nutrients that set the limits of
biomass and production (OECD 1982) but also bring mercury (Håkanson et
al. 1990), lead (Evans and Rigler 1985), PCB’s (Macdonald and Metcalfe
1991, Taylor et al. 1991) and acid (Neary and Dillon 1988). In most lakes,
planktonic primary production is insufficient to meet the energetic demands
of even the plankton, let alone the benthos and fish (del Giorgio and Peters
1993), so energy subsidies are required from the littoral or the catchment. The
modern lake is not a microcosm.
From Forbesian microcosm to empirical holism. Limnologists had elected
to follow what Forbes calls “the a priori road” whereby we begin with bold
conjectures about nature. There is no harm in such a first step. Problems arose
because we did not see this process as only a first step to the identification of
hypotheses for testing. We instead treated some conjectures as inspired
guesses at intelligible principles for ecology. Such principles were accepted
as true. Since they did not need to be tested, they were above the scrutiny of
scientific criticism. With hindsight, we can be more critical; what we see is
that many of the positive attributes of lakes as study sites were illusory. No
measurements showed that the lake was more isolated than a forest or grass-
land, that its inhabitants were more primitive, that its interactions were sim-
88 WHY LIMNOLOGY?

pler, or that its scale was smaller. We had accepted our conjectures as descrip-
tions of reality, rather than hypotheses for testing, and we built and tested
hypotheses within the framework of that preconception. In other words, we
confused our most fundamental theories about the environment with facts.
The change in perception that saw limnology move from the view that
lakes are isolated microcosms to the view that they are part of a larger unit
including their drainage basin and airshed represents a shift in the paradigm
of limnology. Eventually, the old paradigm failed because it could not address
the new problems of pollution, so its initial flaws became more obvious. This
change should entail reassessment of the advantages of lakes for the testing of
ecological theory.
The loss of limnology’s privileged position among ecological testing
grounds also has another, more hopeful message for limnologists. The Forbe-
sian view that lakes were specially appropriate for ecological study because
they were distinct ecosystems was also a trap. The rationale that led us to
expect to see the principles of ecology more clearly in lakes also implies that
the lessons learned in lakes would not easily apply to other ecosystems. This
limitation was not a problem because limnologists found few principles and
because the premise that all ecosystems were similar was not treated as a
theory, but as a metaphysical doctrine. In any case, we can now see that lakes
may not differ from other ecosystems, so our limnological experience may
have broader relevance.
It is counterproductive to think of paradigms as right or wrong. For all its
faults, the old Forbesian paradigm played an important role in the evolution of
limnology. For many years, it attracted scholars to work on lakes and it
allowed these researchers to treat lakes in a consistent and characteristic way.
Because the lake was seen as a homogeneous entity, limnologists came to char-
acterize the ecosystem with a few state variables. In the water, we measured
phosphorus concentration, chlorophyll concentration, zooplankton biomass,
conductivity, and pH. In the sediments, we measured organic content, redox
potential, metal levels and density of macrobenthic invertebrates. Such data
represent the measurements made by a whole generation of limnologists.
Regardless of the status of the lake ecosystem concept, we can use past
measurements to construct new theories about lakes. When we do this, we use
the time-honoured approach that scientists have always used: we identify a
system of interest, we look for regularities in the behaviour of the state vari-
ables describing that system, we express this regularity qualitatively, and then
we may seek to explain these patterns.
For example, Richard Vollenweider (1968) sought patterns in lake
eutrophication. He saw a pattern in the loading of nutrients (P and N) to a lake
and the degree of eutrophication, and developed a simple model to describe
89ECOLOGICAL THEORIES

the pattern. This approach has since been used repeatedly to provide simple
empirical descriptions of nature (Peters 1986, Seip and Ibrekk 1988). For a
small group, this process has become a new paradigm, based on empirical
holism and called “ecometrics” (Håkanson 1991) or “predictive ecology”
(Peters 1986; Chapter IX).
If lakes are no more isolated than other ecosystems, there is no reason to
suppose that this approach is limited in its effectiveness to lakes. It should
and does apply to forests (Box 1981), grasslands (East 1984, McNaughton et
al. 1989), seas (Nixon 1988, Iverson 1990) and rivers (Morin and Peters
1988, Hoyer and Canfield 1991). It should apply everywhere we take the
trouble to measure state variables, to examine those measurements for pattern
and to express those patterns quantitatively (e.g. Fig. 21). Since robust pat-
terns beg explanation, the last step to explanatory theory will follow almost
automatically. Limnology continues to give leadership to ecology, because it
has demonstrated that ecology can be a predictive, empirical science.
90 WHY LIMNOLOGY?
Fig. 21. Relation between plant biomass and primary production in terrestrial and
aquatic systems. One regression applies to both the upper panels. Both it and the re-
gression for forest and woodlands are reproduced in the panel for aquatic systems.
(From Peters 1980)
Plant Biomass (t ha
–1
)
Primary Production (t ha
–1
yr
–1
)

Limnology and marine science. I hypothesize that limnology achieved
its current predictive power (e.g. Table 9) because it passed through a Forbe-
sian period whereby different systems were characterized by a few measure-
ments. We have now forsaken the idea that lakes are isolated ecosystems, but
in doing so, we discovered that those measurements have an independent
value. The importance of limnology’s Forbesian past can illustrated by com-
paring the developments in limnology and oceanography.
Oceanographers have typically led limnologists in breaking new ground,
rather than vice versa. For example, in my own fields of interest, oceanogra-
phers developed techniques to measure phosphorus (Atkins 1923), identified
zooplankton feeding as worthy of study (Pütter 1909), and provided the first
effective techniques and estimates of filter-feeding (Gauld 1951). Therefore
if limnology has stolen a step on oceanography, it is not because freshwater
scientists are usually first off the mark.
Differences in the evolution of the two subdisciplines are not imposed by
the nature of their material. Marine systems offer most of the practical advan-
tages as lakes. Their boundaries are distinct, much of the biota consists of
minute organisms in a homogeneous, three-dimensional habitat with little
structure and few gradients. They are sampled in similar ways and similar
measurements are compiled. The open sea even appears more isolated from
91ECOLOGICAL THEORIES
Dependent variable Units Equation r
2
n
[Chlorophyll] mg m
–3
Y = 0.073TP
1.4
0.96 77
Transparency m Y = 9.8TP
–0.28
0.22 87
[Phytoplankton] mg wet wt m
–3
Y = 30TP
1.4
0.88 27
[Nanoplankton] mg wet wt m
–3
Y = 17TP
1.3
0.93 23
[Net plankton] mg wet wt m
–3
Y = 8.7TP
1.7
0.82 23
[Blue-green algae] mg wet wt m
–3
Y = 43TP
0.98
0.71 29
[Bacteria] millions ml
–1
Y = 0.90TP
0.66
0.83 12
[Crustacean plankton] mg dry wt m
–3
Y = 5.7TP
0.91
0.72 49
[Zooplankton] mg wet wt m
–3
Y = 38TP
0.64
0.86 12
[Microzooplankton] mg wet wt m
–3
Y = 17TP
0.71
0.72 12
[Macrozooplankton] mg wet wt m
–3
Y = 20TP
0.65
0.86 12
[Benthos] mg wet wt m
–2
Y = 810TP
0.71
0.48 38
[Fish] mg wet wt m
–2
Y = 590TP
0.71
0.75 18
Average primary prod. mg C m
–3
d
–1
Y = 10TP – 79 0.94 38
Maximum primary prod. mg C m
–3
d
–1
Y = 20TP – 77 0.95 38
Fish yield mg wet wt m
–2
yr
–1
Y = 7.1TP
1.0
0.87 21
Table 9. Selected regressions describing the relationships between total phosphorus
concentration (TP, mg m
–3
) and other lake characteristics; n: number of lakes. (From
Peters 1986)

the land. Of all the ecological sub-disciplines, marine biology seems the most
likely to resemble limnology, yet in some respects the two sub-disciplines are
poles apart.
There are differences between lakes and oceans. The sea is salt, the scale
of the oceans is greater, sampling is more expensive, biological diversity is
higher and so on, but none of these differences seems crucial. A more impor-
tant distinction between lakes and oceans is the number of discrete sites.
There are vast numbers of lakes. My home province of Quebec has a million
lakes larger than 10 ha in area and, next door, Ontario has half a million more.
There may only be seven seas.
The great number and small size of lakes offer the advantage that limnol-
ogists can manipulate lakes to test our theories (Schindler and Fee 1974; Elser
and Carpenter 1988). In the sea, one could use estuaries, bays (Håkanson and
Wallin 1991) and fjords (Aure and Stigebrandt 1990) as natural units for
replicated measurements and experimentation, but this approach has only
proven popular in the marine littoral (Paine 1977).
The large number of lakes is a misleading statistic. That alone need not
impose different frames of reference on fresh and salt water researchers.
There are more lakes than seas, but those many lakes cover less than 1% of
the area of the oceans, contain less than 0.01% of the ocean volume, and are
spread less evenly across the earth’s surface. In some important ways, the
ocean offers more sampling sites than lakes. Even the discreteness of lakes
may be more apparent than real, for most lake districts bind their lakes
together by common patterns of geology, climate, and land use. In any case,
the distinctness of lakes and the inter-connectedness of the seas are relevant
only as properties of our theories about these systems. If the theories do not
work, then the properties are irrelevant.
Because oceanographers could not delimit small distinct areas to study,
many of them adapted a different form of the ecosystem concept. Rather than
characterize each area with a few state variables, they tended to treat each
study area as a unique assemblage that required detailed description of its
components, likely in terms of a simulation model. Oceanographers felt the
collapse of the microcosm view less. They did not normally work at the level
of the ecosystem, but at a lower level of organization, with ecosystem com-
ponents. They experienced neither the discomfort of paradigm change nor the
benefits of a fresh outlook. Instead, marine biologists continued to treat their
material as an aggregate of many sub-systems, each of which has to be indi-
vidually described. This view is discussed in the next chapter. For the present,
it is enough to note that, without the Forbesian frame of reference, marine
ecologists could not see their data as state variables characterizing a system,
and therefore their systems remained difficult to characterize or to compare.
92 WHY LIMNOLOGY?

Why limnology — an answer. In summary, limnology originally played
a leading role in ecology because limnological systems seemed optimal for
testing contemporary ecological theory. The suggestion that the limnetic
pelagial might be a microcosm attracted some of the brightest minds in ecol-
ogy to consider limnological examples. Terrestrial ecologists were less drawn
to the approach because their ecosystems were less clearly connected inter-
nally; marine biologists were not attracted because their potential ecosystems
were less obviously isolated from other parts of the ocean. Neither group was
able to agree on how to identify the system they should study or on what
measurements characterized their systems. Confident in the definition of their
ecosystems, limnologists moved ahead to describe their ecosystem with state
variables that characterized the whole.
Subsequent development revealed anomalies and flaws in the limnological
paradigm, and many of the concepts it encouraged have been shown to be
untestable or erroneous. If limnology is no longer leading all of ecological
theory, it is because its earlier success was premised on the misapprehension
that tests of ecological theory required an isolated microcosm. Since this is no
longer tenable, ecological theories are equally testable in many systems and
limnology has lost some pride of place. Thus to much of ecology, limnology
seems to have slipped from its once pivotal position (Jumars 1990, Wetzel
1991).
Some limnologists learned from the experience to move beyond the micro-
cosm concept. The information and tools that developed under the old
paradigm allowed researchers to step boldly into a new phase of growth based
on measurable state variables and prediction of whole system properties.
Although our original conceptions of lake system limits were flawed, and
although we are uncertain what the new limits should be, the holistic mea-
surements we took still describe our lakes. Even if we can no longer define
the system, our holistic measurements function as the basis of empirical theo-
ries to predict lake characteristics.
Science does not depend on what we think about reality or about the state
variables we use. Science depends on how well these variables function in
theories to make predictions. Some limnologists learned this when their
paradigmatic conception of reality crumbled, but related empirical theories
continued to predict. Predictive limnology is a flourishing sub-discipline
(Peters 1986, Seip and Ibrekk 1988), and limnology is once again showing
the way, this time towards a new science of predictive ecology. And that is
why we should teach and study limnology.
93ECOLOGICAL THEORIES


VII Reductionism versus Holism:
An Old Problem Rejuvenated by the Computer
“The meaning of this illumination [...] did not
penetrate at once, and notably the word trim [...]
long remained obscure.”
Samuel Beckett
[Molloy (1950)]
The subject of this chapter has been ignored for such a long time that the
key words in the title are nearly meaningless to most biologists. These
words may even give the impression that I will deal with the history or
philosophy of biology rather than with its science. That is not my inten-
tion at all, for I am neither an historian nor a philosopher. My only rea-
son for being interested in the history and philosophy of science is that
these disciplines may teach me to do my science, ecology, more effec-
tively.
Ecology is a branch of science that is, or at least ought to be, very impor-
tant to society. We need ecological predictions and we need them now, yet
ecology has failed to produce the predictive theories we need. If asked why,
most ecologists explain away our failure by the extreme complexity of our
subject matter and the youth of our discipline. I do not deny these explana-
tions. They may be right. However, the intractability of our subject matter is
not the only barrier to ecological progress. There is also something peculiarly
unproductive about the approach we ecologists have taken to study our sub-
ject matter.
The Place of Philosophical Debates in Biology
The theme of this chapter is that we ecologists have been working ineffi-
ciently because we ignored philosophical debates that involved a few biolo-
gists a century ago. We working scientists ignored the biophilosophers
because we never understood what they were talking about. Indeed, they did
not understand it themselves. The advent of the computer has re-invigorated

some of these old, semi-philosophical questions, and forced us to restate them
in such a way that they become meaningful to experimental ecologists. I will
illustrate the problems this creates with some of my own work with ecosys-
tem ecology in the high Arctic and with the autecology of zooplankton feed-
ing. Finally I will suggest that if we learn the lesson such examples provide,
we may become more effective. The point of the chapter is to suggest that the
discovery of ecological systems and the introduction of the technique of sys-
tems analysis into biology require us to reevaluate the ecologists’ approach to
science.
Molloy and the principle of trim. Before describing the history of our
debates I will give an example of what I mean by saying that we did not
understand our own arguments. This statement would seem strange to
philosophers and historians because they work with words, and are expected
to use them rigorously. However, scientists tend to be less interested in
words than in phenomena. Consequently, when a scientist looking at nature
gets a brilliant idea, the immediate problem is to express the idea in words.
Until this is done, there will be no convincing others of the brilliance of the
idea.
My illustration comes from the writings of Samuel Beckett — a writer
most remembered for his deep understanding of the human tragedy. I believe
he had an equally deep understanding of science, as shown beautifully by the
incident of the sucking stones in his novel Molloy, which forms the prologue
to this book.
Molloy, crippled, destitute and lost, rests (to the extent that he could ever
rest) on the sea shore. There he sucks wave-washed pebbles to relieve the
pangs of hunger and thirst. Yet he is totally engrossed in the problem of dis-
tributing his sixteen sucking stones among his four pockets such that he can
suck them each in rotation. He goes through several unsatisfactory solutions
until finally he has an insight. He can obtain his goal if he sacrifices “the prin-
ciple of trim” and distributes the stones unevenly among his pockets. At this
phase in his research, Molloy exemplifies my point, that scientists have trou-
ble expressing their discoveries, in the quotation that begins this chapter.
Molloy was convinced he had made a great discovery and he expressed his
discovery in a word: “trim”. However, for a long time he did not know what
he had discovered, nor why it was aptly described as “sacrificing the princi-
ple of trim”.
This inadequacy did not deter Molloy. Neither does it deter the biologist
who has found something great but cannot explain it. This I believe was the
situation with a few biologists who took part in the debates of the 19th cen-
tury. Their insights were important, but unfortunately the words they used left
the great mass of working biologists unmoved.
96 REDUCTIONISM VERSUS HOLISM

Vitalism and mechanism. The debate I want to discuss is the debate
between the holists and the reductionists. Because this debate has become
incredibly confused, I will also give some of its history and show how it
evolved.
Aristotle is again a good starting place. Among his many ideas about the
nature of living things, he believed that the difference between living and
non-living things was due entirely to the fact that the living things possess a
soul. A simple consequence of this belief is that we cannot expect to learn
anything about life by studying a dead body. Neither can we learn anything
about life by studying a hand or a liver or some other organ removed from a
living body. This philosophy (perhaps “faith” is a better word) was called
vitalism (Table 10).
Vitalism dominated biology until the 18th and 19th centuries, by which
time physiologists had wrought a slow, initially unnoticed, change. Incited by
René Descartes (1596–1650), they began to treat living things as if they were
physico-chemical machines. For example, Hermann Boerhaave (1668–1738)
and George Martine (1702–1741) argued that the vital warmth of the human
body is produced by the friction of red corpuscles rubbing on the walls of
arterioles. A century later, Gustav Magnus conducted experiments with blood
removed from the body, analyzing the results of his experiments as if blood
were merely a physical fluid (Mendelsohn 1964). Theodor Schwann
(1810–1882) extracted the active principle of gastric juice from the lining of
the stomach with acid, and then showed it could be precipitated as if it were
simply a chemical substance (Bodenheimer 1953).
97THE PLACE OF PHILOSOPHICAL DEBATES IN BIOLOGY
Vitalism: Life depends on the presence of a soul.
Mechanism: Living organisms are simply complex physico-chemical machines.
Organicism: Life depends on organisation; each level of increasing complexity of
organisation can only be described in terms of laws appropriate to
that level.
Reductionism: The proper approach to the study of complex phenomena, like life, is
to decompose this complexity to simple components. In many cases,
these components can then be shown to be instances of general phys-
ical and chemical laws.
Holism: Complex systems must be treated as whole systems, because the
process of analysis inherent in reductionism destroys the basis of
their integrity.
Table 10. Fundamentally different approaches to the study of biology

The story of vitalism ends in France in the middle of the 19th century
in a little cellar in Paris, where the great physiologist Claude Bernard
(1813–1878) had a table which he called his laboratory. Bernard did a very
simple experiment. He killed a dog and removed its liver. He then inserted a
tube into the hepatic portal vein and perfused water through the liver. He ana-
lyzed the water emerging from the liver for sugar and found glucose, but in
time no more sugar came out. He had washed out all the available sugar.
He then left the liver in a warm place for several hours, once again perfused
it, and found more sugar. Bernard concluded that the liver could make
sugar from another compound and he called this hypothetical precursor
“glycogen”.
This experiment was terribly important in its day. It showed that animals
can change one compound into another, it helped Bernard develop the idea of
internal control, which became one of his greatest contributions to science,
and it showed that an organ can have more than one function (Larner 1967).
The experiment was even more important because biologists of the time
accepted it as valid.
Bernard had drawn conclusions about the functioning of a living organism
from an experiment done with a part of that organism separated from the rest
of the body. For the two thousand years after Aristotle, this approach had
been totally unacceptable to most biologists. Everyone had known that the
only difference between a living organism and a dead organism is the soul,
which conferred life and left the body at death. Thus a dead animal or animal
part could tell us nothing about life.
By Bernard’s time, biologists were no longer unanimous on this point. A
growing number were treating organisms as highly complicated machines.
The biologists promoting this new approach were the mechanists who saw
life as a complicated physico-chemical process, and Bernard’s experiment
marked the triumph of this scientific revolution. The real significance of
Claude Bernard’s experiment was that it was the last shot in the battle. With
this experiment, the mechanists won the war and vitalism was never again an
acceptable paradigm for biologists. With Claude Bernard, the revolution was
over and most biologists rejected vitalism for mechanism.
Organicism and holism. Our story does not end with the mechanistic
revolution. It begins there. Some biologists accepted that biology could best
progress without postulating a soul, but not that living things were only per-
ambulating bags of chemicals. These biologists argued that the components
of living things are organized into living systems, and that the properties of
these systems could not be predicted from a study of their parts. This new idea
was a modification of vitalist thought. It differed from vitalism by substitut-
ing organization for soul and by asserting that we could learn to understand
98 REDUCTIONISM VERSUS HOLISM

living things, but only if we studied them intact. This view came to be called
organicism. The organicists produced the idea that is central to this chapter.
The new trend began with Xavier Bichat (1771–1802) who was obsessed
by the variability of living systems. Physico-chemical systems were depend-
able and their behaviour reproducible, but biological material was unreliable
and variable. From this hypothesis, he argued that biological systems must
have something extra. Their behaviour must be governed by new, still
unknown, peculiarly biological laws (Mendelsohn 1964, Hall 1969).
Lloyd Morgan (1923) improved on Bichat by inventing the idea of “emer-
gent properties” whereby each level of complexity has its own laws. An
important consequence of this idea is that if we want to develop theories
about matter at a particular level of complexity — like a cell, an organ or a
whole organism — then we must study the cell, the organ or the whole organ-
ism, not its parts. This is because the whole cell (or organ or organism) has
properties that its component parts lack.
At the beginning of the 20th century, the opposing forces had regrouped
and the mechanists had changed their names. The descendants of mechanism
became “reductionists”, because they believed in breaking up the system they
intended to study. The battles that raged, unnoticed by most biologists, were
first between the vitalists and mechanists and then between the mechanist-
reductionists and organicists. It was the organicists who had something to say
but were unable to find a way of saying it. Just as “trim” meant something to
Molloy, “emergent properties” meant something to the organicists. Unfortu-
nately, it meant nothing to most working biologists.
In the 20th century, their words began to gather more meaning and began
to be applied not just to living organisms but to ecological work as well. The
first advance was made by Ludwig von Bertalanffy (1950, 1952) who sug-
gested why living systems might have emergent properties. Von Bertalanffy
said the important characteristic of organisms is that they are systems, that is
to say they comprise “a complex of elements in mutual interaction”. The
behaviour of a system depends on all interactions amongst all the parts, and
so the interactions are as important as the parts. If one part is removed, the
interactions between it and other parts are broken. This was much more con-
vincing than the vague talk about emergent properties, and in recognition of
the advance we changed the names of the protagonists for the last time to
holists and reductionists.
Reductionists and holists were debating a real and significant question
about scientific knowledge, but we hear little about their dispute today. I
believe biologists lost interest because the opposing forces were just too
unequal to make a battle worth fighting. The holists were few and far
between, so the reductionists just ignored them. Consequently, most of us are
99THE PLACE OF PHILOSOPHICAL DEBATES IN BIOLOGY

reductionists by default. We accepted an approach to which we were condi-
tioned without ever questioning our belief or knowing that there is another
viewpoint.
Holism and reductionism in ecology. Most ecologists were oblivious of
these questions. We were mostly concerned with predicting and controlling
the abundance of particular species, because we wanted to know, for example,
if fishing would deplete fish-stocks, or how different forms of pollution affect
organisms we care about. Our interests were therefore largely directed to
individual species, and we hoped that a single factor would be of overwhelm-
ing importance for each species.
Many ecologists behaved as if the distribution and abundance of a species
could be predicted from physical and chemical factors alone, and early field
work was largely directed at the relations between individual members of a
species and their physical or chemical environment. Researchers therefore
sought to establish the response of an organism to temperature, humidity, pH,
salinity, etc. Given this emphasis, it is scarcely surprising that autecology was
a major branch of ecology or that the limiting factor was the most popular
concept. The emphasis on autecology even allowed plant and animal ecology
to develop as separate disciplines in different university departments, as if
animals and plants did not interact.
The autecological approach was appealingly simple, but it did not work
well enough. Researchers therefore increasingly emphasized biological inter-
actions, like competition and predation. This interest marked an implicit
recognition that biological interactions can be more important than physico-
chemical factors.
The American botanist F. E. Clements (1916) moved decisively beyond
autecology when he wrote about plant communities as superorganisms. He
has since been thoroughly abused for this. I think the attacks against him were
unjust, but I recognize that he made two fatal mistakes: he was suffering from
the Molloy syndrome and he was too far ahead of his time. If Clements had
“penetrated the meaning of his illumination” and if he had read von Berta-
lanffy, he would have said something like this: “The community is a system,
and in this respect, it resembles an organism. But it is more complex. It com-
prises the interactions among many organisms, and in that sense, the commu-
nity is at a higher level of organization than an organism. This is what I mean
by superorganism”. Similar ideas are currently accepted as elements in hier-
archical thinking (Allen and Starr 1982, O’Neil et al. 1986). But, alas for
Clements, he did not say it right, and he has been a bogey-man for politically
correct ecologists ever since.
Nevertheless, gradually and unconsciously, we began to change. We ad-
mitted the food chain among our fundamental concepts, and when faced with
100 REDUCTIONISM VERSUS HOLISM

results like Hardy’s (1924) study of the complex feeding relations of the her-
ring, we admitted that the food chain was unrealistic and changed to food
webs, implicitly accepting the idea of multiple interactions. We stopped sep-
arating plant and animal communities, and began to talk about ecosystems.
We slowly came to realize that in ecology we are not dealing with isolated
entities but with one or more highly organized systems in which the parts
interact with and depend upon one another.
The concept of the ecosystem led to a problem. Most ecologists had devel-
oped a tradition of studying bits and pieces, but had no idea of how to fit our
bits and pieces together. Then, two developments outside of ecology made it
possible for us to deal with complex systems. These were systems analysis
and the computer.
What is Systems Analysis?
I can explain systems analysis best with a simple example (Fig. 22). Take
a tumbler of water containing plant nutrients. Shine a light on it. Put in some
algal cells and add a few water fleas to eat the algae. This is a system: algae,
nutrients and Daphnia all living together and dependent on each other. If we
know the quantity of each component and the interactions between all the
101WHAT IS SYSTEMS ANALYSIS?
Fig. 22. A simple system consisting of an external energy source (light), a primary pro-
ducer (algal cells), herbivores (Daphnia) and their interactions

components, then we can use a computer to do an incredible amount of sim-
ple arithmetic and so calculate the amount of each component or “state of the
system” at any time in the future. This approach was taken by many ecolo-
gists who accepted that ecology deals with complex systems. It is called
“systems” analysis because it seeks to decompose the working natural system
into parts for study, and then to recreate that system as the sum of these inter-
actions. In the example of Fig. 22, the interactions I must measure are feeding
rate as a function of animal size and algal abundance, uptake rate of plant
nutrients as a function of nutrient concentration, and the rate of nutrient
excretion by the animals.
There is a catch. To succeed, I must measure all components and all inter-
actions. If I leave out Daphnia or omit excretion, or if I measure one interac-
tion erroneously, my model will not work any better than a watch without its
tiniest cog-wheel. To do otherwise, to presume that we can either leave some
elements out of our model or model only the significant interactions, presup-
poses that we can somehow identify the important processes a priori.
Failures since the time of Aristotle should have taught us that appeals to
“intelligible principles” or the “a priori road” of assumptions about nature are
not effective. If we are to dismiss some interactions as unimportant, we must
first study those interactions in detail. Unfortunately, that effort is exactly
what we are trying to avoid. Moreover, partial representations involve a pos-
sible contradiction because the appeal of mechanism and reductionism is
their capacity to represent “what’s really happening”, so that we will “under-
stand what’s going on”. If we ignore much of what is really happening, then
the apparent strong point of the reductionist-mechanist position is already
lost. Any argument would only be a methodological debate about how thor-
ough the holist position should be.
Even in a simple system, the possibility of modelling all the interactions is
small. For example, we perhaps should consider the excretion of different ele-
ments or compounds separately. Perhaps our model should have a place for
the bacteria in the daphnid’s gut and the epibiotic organisms living on its
shell. Perhaps the modifying effects of time of day, or light levels or temper-
ature or age of the culture should be considered. Perhaps there are important
interactions among the algal cells. Neither these specific problems nor the
general question of when we should stop analyzing the system has been
resolved.
We are not satisfied with systems consisting of a few species and their
physico-chemical environment. We need to describe large ecosystems, and to
do so we must gather larger and larger teams of scientists and technicians to
work on each ecosystem. For example, in Russia, a team of 150 scientists
worked on a single reservoir for three years. This trend to more ambitious
102 REDUCTIONISM VERSUS HOLISM

projects is also exemplified by large scale international programs, like I.B.P.
(The International Biological Programme) of 1965 to 1975, or the contempo-
rary JGOFS (The Joint Global Ocean Flux Study) and WOCE (World Oceans
Circulation Experiment).
To appreciate the difficulties that systems analysis presents in the treat-
ment of these large systems, we must consider the data that ecologists are
gathering and the uses to which these data are put. Imagine the moderately
simple ecosystem represented by the biota of a temperate lake. In round num-
bers, this might include 200 species of plants, 100 species of herbivores, 100
species of carnivores, 50 species of bacteria, 200 animal species that cannot
be attributed to any trophic level, and 100 species of detritivores. This com-
munity of 750 species is simpler than it might be. Likens (1992) estimates
that there are at least 850 species in Mirror Lake, New Hampshire (USA) and
Elton (1966) suggested that Whytham Woods in England might contain 5000
species of metazoans. To model such systems we must describe these com-
munities. Such a description might take the form of a compilation of the
biomass, the rates of birth, growth, and death for each species, and the inter-
actions (predation, competition, etc.) among these species. We might further
describe the effect of variations in light, temperature, nutrients, etc. on these
components and their interactions. We could then build these parts of the sys-
tem into a model system in our mind (Fig. 23, overleaf).
The arrows in Fig. 23 show only the predator-prey interactions. In a real
description of the system, every component potentially interacts with every
other one. Therefore if the number of components (n) equals the number of
species, n = 750 and the potential number of interactions = n(n–1). This aver-
age lake has 561 750 potential interactions among its components. (The cal-
culation assumes that the effect of Species A on Species B must be described
separately from the effect of Species B on Species A, because that situation
seems more plausible to me; if the interaction is instead bidirectional, the
number of arrows will be halved.) Now imagine that we have our system
arranged in our minds where all 561 750 interactions are indicated by arrows.
The human mind cannot keep track of all these interactions. To predict the
effect of any disturbance on our system, we have to use the technique of sys-
tems analysis and its characteristic tool, the computer. What should we do
next to make this possible?
There is no need for details. It is sufficient to note that systems analysis
requires us to quantify every interaction, under the entire range of environ-
mental conditions that we expect to encounter. If each interaction requires
only one person-year of effort, this description will require centuries even for
a large team. In passing, I note that I spent over four years developing a
theory that describes only 50% of the variation in a part of a single inter-
103WHAT IS SYSTEMS ANALYSIS?

action: phosphorus excretion rate by Daphnia rosea (Peters 1972, Peters and
Rigler 1973). Other arrows linking zooplankton, their food and the resources
upon which that food depends were studied almost as long with almost as
limited success by other doctoral candidates in the same laboratory (Cham-
berlain 1968, Confer 1969, 1972, Haney 1970, Lean 1973). I conclude that
full description of an entire ecosystem is so great an effort that we will never
complete the task. In fact, we won’t even try. The job is just too big.
Some problems with proposed solutions. The previous section estab-
lished that it is impossible to specify the current state and interactions of the
components of complex ecological systems. The problem is not a philosoph-
ical issue but a purely practical limitation (Wimsatt 1980). Simplification
seems a practical solution, but a few moments’ reflection shows that this is
not the case.
Identifying the components. Most biologists prefer to work on individual
species, and when systems models have been built, they normally depend
104 REDUCTIONISM VERSUS HOLISM
Fig. 23. A hypothetical lake ecosystem consisting of 200 species of primary producers,
100 species of herbivores and 100 species of carnivores. Arrows indicate potential
trophic interactions, and not shown are 50 species of bacteria, 100 species of detriti-
vores, 200 species that could not be classified to a single trophic level, and all non-
predatory interactions

heavily on autecological work done at the species level. Unfortunately, we
do not know the number of species in any natural ecosystem. Study after
study has shown that the harder and longer one searches, the more species
one will find. Thus we can never know how many components comprise the
system.
One can seem to address this problem in a number of arbitrary ways. For
example, one could propose that the system is characterized well enough
when the rate of increase in species number per unit of additional effort falls
below some specified value. Unfortunately, approximations like that will not
do. There are many cases in which obscure components play important roles
in structuring their communities: Opuntia, the prickly pear cactus, is held in
check by the now rare Cactoblastis moth in Australia; in Africa, intestinal
parasites of tse-tse flies apparently protect the Serengeti from over-exploita-
tion by debilitating human agriculturalists with sleeping sickness. Currently
small or rare members of the ecosystem cannot be excluded from a systems
analysis on the grounds that such components never play an important role.
Aggregation. One way to simplify the system is to lump different species
of similar organisms into a smaller, more tractable number of functional
groups. The trophic level approach (Lindeman 1942) is one such attempt. The
description of the planktonic community of primary producers in terms of
chlorophyll concentration might be another. Unfortunately, aggregation is
rarely easy or effective in systems analysis. To determine which organisms
are functionally similar members of a community, we must determine their
relations to the rest of the community, but this was the difficulty we were try-
ing to escape. Whether we aggregate the data or not, we are required to create
a tiny theory for each member of our system in which we hypothesize how
that member relates to every other component. Since we can hardly expect to
create 561 750 correct theories, we must build error and approximations into
the systems analysis model. Since these errors will be propagated with each
calculation (Beck and Halfon 1991, Peters 1991a, van Straten and Keesman
1991), we cannot expect that the model will work.
Interactions. We have not seen it all yet. One last problem poses even
more difficulties than either simplification or aggregation. Consider how the
ecologist usually quantifies interactions between system components. Occa-
sionally these are estimated in situ, but more often than not this is impossible.
Therefore the two interacting components to be studied are isolated and their
interactions are measured in the laboratory. For example, when we measure
the feeding rate of a water flea as a function of the abundance of its food, we
isolate the flea and one type of food in a beaker where we can easily vary the
food supply. Then we take the rates we measured and apply them in our
model.
105WHAT IS SYSTEMS ANALYSIS?

This procedure is effective only if no component of the natural system acts
directly on the interaction we are measuring. If there is such a component, our
lab values will not apply to the natural system. Obviously such modifying
interactions are common: the presence of noxious algae may cause the water
flea to reduce its ingestion rate of palatable species (Burns and Rigler 1967,
Burns 1968), the absence of vitamins may make the water flea less capable of
utilizing assimilated food (Provasoli et al. 1970), and feeding rates change
with temperature and time of day (Haney and Hall 1975). Since most interac-
tions are modified by the environment in which they occur, we must study the
animals in situ, and we must study each interaction intensively. We cannot
escape the massive work-load that systems analysis of any natural ecosystems
imposes (Rigler 1982a). But since we cannot do that much work, the applica-
bility of systems analysis is so limited as to require reassessment of the
approach.
Two Personal Experiences
The impossibility of really analyzing a system and of synthesizing the
parts into a computer model is rarely appreciated. Most of our research is
designed to allow us to analyze the bits of a natural system that interest us,
leaving the remainder for others and assuming that someone else will solve
the problems of putting the big picture together. I no longer accept this view
because I have seen the magnitude of the problem of creating a system anal-
ysis model. I encountered these difficulties in many aspects of limnology,
but I will illustrate the problems with two aspects of my work which dif-
fered very much in scale: the Char Lake Project and the analysis of zoo-
plankton feeding.
The Char Lake Project. Some years ago I became involved in the study
of one of the simplest lake ecosystems in the world, Char Lake located on
Cornwallis Island in the Canadian High Arctic. This study was not under-
taken because the lake seemed an ideal site for systems analysis, but we
justified our research with a conceptually similar argument. We hoped that we
could exploit the simplicity of Arctic systems (Table 11) to increase our
chances of learning something about lakes in general. In retrospect, this
justification seems lame, but at the time it was quite convincing.
The Char Lake Project also represented part of Canada’s contribution to
the International Biological Programme in which 50 countries participated in
over 140 different projects. Each project involved a team of biologists work-
ing together for up to five years. At the time, IBP provided an unprecedented
level of funding, international communication and communality of purpose.
106 REDUCTIONISM VERSUS HOLISM

In many ways and despite its eventual shortcomings, the period of the IBP
was a golden age in ecological research.
The ostensible reason for the IBP was to determine the limits of produc-
tivity of the earth and thus to determine its capacity to meet human needs.
Teams of scientists were to measure productivity all over the earth, and the
composite of their results was to represent this global overview. Most of us
felt that this was too simple a goal and sought to measure primary, secondary
and tertiary production and the associated biomasses and energy flows as
well. We hoped that by focusing our attention on this previously little studied
set of properties, and by standardizing our techniques as much as possible, we
might discover a theory relating these properties to others such as light, tem-
perature or nutrients. This hope now seems naive.
Regardless of the reality of our hopes, the same data were to be used both
to estimate production and to try to understand production processes. A large
team was required for this work because the objective was to study every-
thing in the system.
Only a few members of the IBP were directly involved in simulation mod-
elling. The others sought to identify the components of the system, and then
to quantify each component and interaction. When these had been measured
in sufficient detail, the various components were to be pieced together as a
predictive model in a computer simulation.
There are reasons for comprehensive data collection, other than systems
analysis. Indeed, the larger part of the IBP teams doubted that systems analy-
sis would resolve the problems of ecosystem ecology. That group acted as
though ecology (and presumably science) consists in the organized and sys-
tematic collection of data that the scientific community considers important.
Presumably few members of IBP were pure Baconians, subscribing to the dis-
credited view that theories arise automatically from a sufficient quantity of
107TWO PERSONAL EXPERIENCES
Group Number of species
Char Lake A temperate lake
Vertebrates 1 20
Planktonic crustaceans 1 10
Planktonic rotifers 1 15
Diptera 7 150
Nematodes 24 ?
Other benthos 13 100
Table 11. The depauperate fauna of a high Arctic lake, Char Lake, relative to that
typical of a temperate lake

good data. However, many would have accepted the view that the collection
of good data is justification enough.
Other researchers may have joined a team project because they believed
that they were working on the single critical and tractable component of the
ecosystem. The subject of their interest would repay study because, when this
component was understood, we would be able to predict the observations that
other members of the team had conveniently made (Fig. 24). A corollary of
this egocentric view is that the subjects studied by others would prove less
easy or valuable. The weak point in this argument is the lack of consensus
about which process was the controller, and the individual assurance that each
of us had chosen to study the critical interaction whether that be nutrient load,
primary production, excretion, predation, or whatever.
Regardless of its rationale, the team approach made reductionists of us all,
even if we did not know the term, because the team was trying to assess the
108 REDUCTIONISM VERSUS HOLISM
Fig. 24. A subjective view of the interactions among lake processes studied by different
reductionists in the same team. (From Rigler 1975a)

whole, but the individuals were studying the parts. The approach also made us
all systems analysts, whether we touched a computer or not, because we
could not hope to put all our studies together except in some form of com-
puter model.
Problems in the components of reductionism. The problems of building a
computer model for an ecosystem are rarely apparent to anyone except the
modellers. The present generation of models is so complex that a description
of all the adjustments, guesses and theories that went into model development
cannot be given in the space of a scientific paper. As a result, these presenta-
tions are usually impossible to assess (Rigler 1976). Instead, I will simply
describe some of the results from our own work that made the simulation of
Char Lake impossible.
So far, I have written as though the species was the smallest component
that one would include in a systems model. However this is a gross over-sim-
plification. For example, although all the fish in Char Lake are Arctic char
(Salvelinus alpinus), these animals are not all equivalent. A few were canni-
balistic giants, whereas the rest of the population fed largely on benthos. The
single crustacean zooplankter, Limnocalanus macrurus, has 11 different
instars or developmental stages, some of which are so different that they were
once classified as a separate genus, all of which differ in their feeding capac-
ities. Thus the complexity of the system is not necessarily reflected in its
species list.
If we could identify the components of the ecosystem, we would then seek
to estimate their biomass. Such estimates have preoccupied ecologists for
many years. Most of us recognize that the size of a wild population can rarely
be estimated within 50% at any one point in time and that population size nor-
mally fluctuates by an order of magnitude over time (Connell and Sousa
1983, Peters and Wassenberg 1983, Schoener 1985, Pimm and Redfearn
1988). For example, when we used nets to make a capture-recapture estimate
of the fish population in Char lake, we estimated a population of 15 000 char.
However, when we counted the fish on the spawning beds, our estimate rose
to 50 000. Apparently, marked fish were more easily recaptured than
unmarked fish, so our recapture rate was high and our population estimate
low. Although we are justified in claiming that the initial estimate was low,
we cannot assume that our spawner count was right.
Finally, our modelling, whether conceptual or computed, required esti-
mates of the various rates of energy flow through the members of the com-
munity. In a general sense, this was unsatisfactory because each component
was analyzed to a different degree and with different methods. A great deal of
effort was directed to the zooplankton, largely because the plankton were the
simplest part of this simple system and because many of us were primarily
109TWO PERSONAL EXPERIENCES

planktologists. Unfortunately, most of the energy flow in this system went
through the benthos, which was less studied and more difficult. For example,
none of the 24 species of nematode could be cultured and there were no in situ
methods for these organisms. The energy budgets of our most speciose taxon
could not be studied.
Some holistic successes within the Char Lake Project. These many diffi-
culties should not obscure the successes of the project, but because most have
been reviewed elsewhere (Rigler 1978), I will only mention some examples
here. At the level of the whole system, Welch (1974) was able to measure
community metabolism by using the ice-covered lake like a huge dark bottle
through the dark Arctic winter. Welch and Kalff (1974) developed techniques
to measure bryophyte-dominated benthic production in the lake and showed
that 80% of all primary production in this lake was benthic. They also showed
that nutrients limited algal biomass in the Arctic, just as they do in more tem-
perate regions (Kalff and Welch 1974). At a smaller scale, work with rotifers
(Rigler et al. 1974), chironomids (Welch 1976), Mysis (Lasenby and Lang-
ford 1972) and char (Holeton 1973) showed that these Arctic populations
were not especially well acclimated to their cold environment, but behaved
like southern representatives of the same species at the same temperature; this
finding is a major challenge to a generation of autecology. With respect to
zooplankton feeding, careful examination of the guts of the crustacean zoo-
plankton showed that these calanoids did not digest diatoms, unlike their
marine counterparts (Kibby and Rigler 1973). Careful use of well-designed
sediment traps showed that Limnocalanus produces only one faecal pellet per
instar; contrary to general belief, the faeces of this animal are not bound into
a pellet and so most of the faeces might not have been lost from the water
column, but rather resuspended.
The common thread through these results is that all address different sys-
tems of interest. The lake, the benthos, the animals, and the faecal pellet were
treated as separate systems, albeit systems of very different scales. Thus suc-
cessful projects focused on a phenomenon for its own sake and not because
the phenomenon was the sum of its component processes or because the pro-
cess played a role in a larger system too. Holistic analyses by individuals or
small teams succeeded, whereas systems analysis of the entire project based
on reducing the whole to parts that were studied separately failed.
Zooplankton feeding. My experience with zooplankton feeding seems
poles apart from the scale of the Char Lake Project. However, decades of
research into this single element in the systems analysis of any lake show con-
clusively that estimations of the parameters for transfers among different lake
compartments are uncertain. I can illustrate this point best with research on
zooplankton feeding, because I know that work well. Indeed, I believe that
110 REDUCTIONISM VERSUS HOLISM

zooplankton feeding rate estimates are among the best measured of all
ecological rates (Rigler 1971, Peters 1984). Unfortunately, even the best
estimates leave much to be desired as system parameters.
Individual variation. If one measures the rates of ingestion of a number of
animals, whether these be Limnocalanus in Char Lake or Daphnia in the lab,
one finds that the animals do not all behave in the same way. Some eat at a
very high rate, others at a much lower rate, and others are in between (Fig. 25;
Turner et al. 1993). When we try to describe these data we often use the aver-
age rate, assuming that our experimental conditions captured nature effec-
tively and that the most representative rates are somewhere in the middle of
our measurements. Alternatively, we could decide that the experiment some-
times disturbs the animals, and that the correct rate is the maximum observed
in the experiments. There is however convincing evidence that some experi-
mental protocols starve the animals before feeding, and that starving animals
feed at greater than normal rates. Perhaps the low values are most appropri-
ate. In any case, we do not know the rates at which animals feed, so the rates
we use in our simulations are hypothetical and uncertain.
Environmental factors and their description. There are other problems.
Early work on zooplankton assumed that they filtered food from a constant
volume of water per unit time. If this was so, filtering rate would have been a
constant, and ingestion rate would rise directly with food concentration
(Gauld 1951). Subsequent studies showed that filtering rate was constant
111TWO PERSONAL EXPERIENCES
Fig. 25. The effect of size on filtering rate in laboratory study of Daphnia. (Peters un-
publ. data)
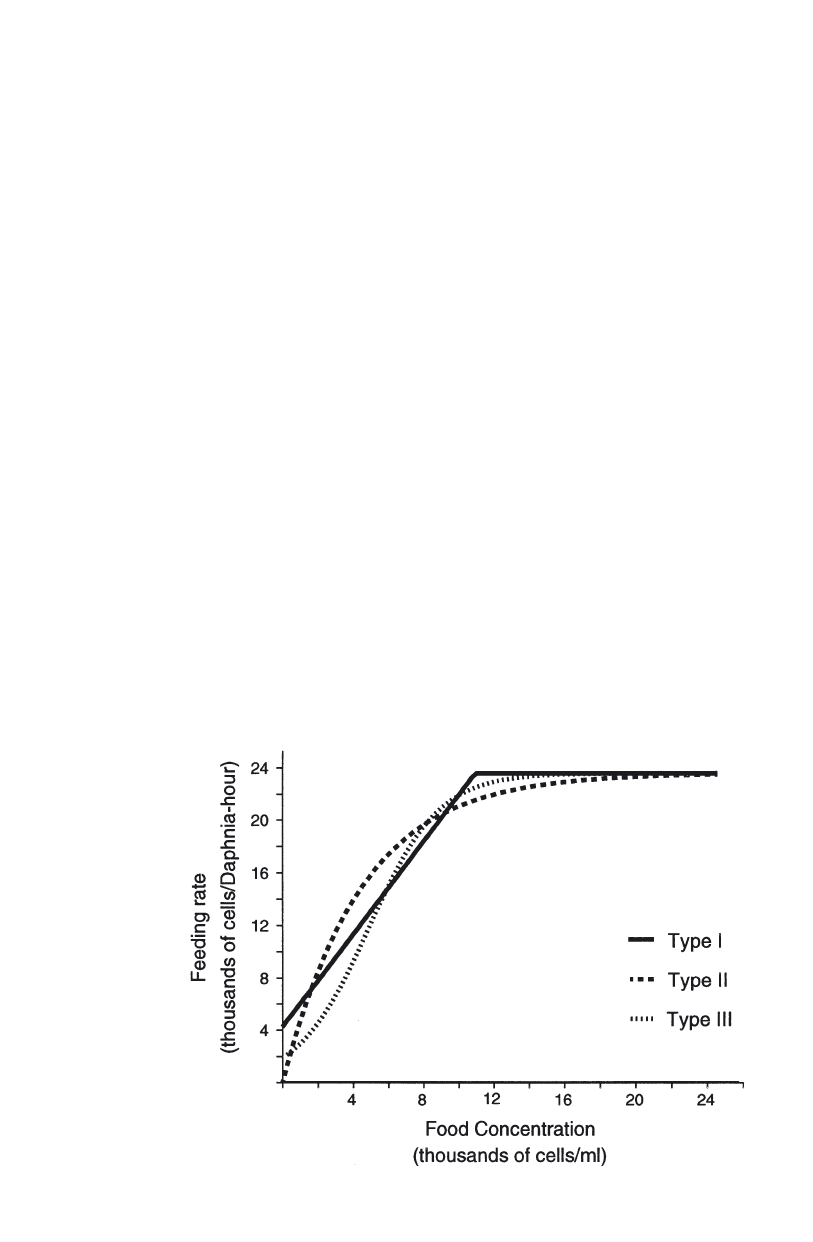
only at low food levels (McMahon and Rigler 1965) and declined at increas-
ingly higher food concentrations. Ingestion rate is instead roughly constant
when food is abundant, and varies with food levels when they are low. Many
different curves can relate ingestion rate to available food, but three are pre-
ferred (Fig. 26) because they relate to three basic types of predator functional
response identified by Holling (1959). The differences among these curves
are critical to some systems analysis models, but the data for zooplankton
have never been good enough to distinguish which model is best. Indeed,
where enough data and enough statistics have been brought to bear on the
problem, the best fit can support any of these three curves, and more besides
(Downing 1981).
Lab measures and field applications. Even if zooplankton feeding in the
laboratory could be effectively described, application of the description to
nature would still be problematical. For example, Haney (1973, Haney and
Hall 1975) has shown that in nature animals follow a marked diurnal cycle in
feeding rate, and approach rates measured in the lab only around dusk and
dawn; so our lab rates may be even poorer approximations of field behaviour
than we all fear.
Still another source of error is the use of relations and measurements in the
literature to convert measured lengths and counts into estimates of population
biomass. Schmidt (1968) has shown that variations in food level can change
112 REDUCTIONISM VERSUS HOLISM
Fig. 26. The effect of food concentration on ingestion rate of Daphnia. (From Porter et
al. 1982)

an animal’s weight two-fold, and many of the discrepancies in estimates of
weight specific rates in the literature might represent no more than the error
due to an inappropriate length-weight relation. I once surprised myself by
generating negative weights for Daphnia simply by “correcting” my mea-
sured weights for the egg mass, in turn estimated as the product of egg num-
ber and an average value for egg weight. Egg size can vary considerably with
age and food level (Glazier 1992), so no such easy step is justified.
Other organisms. One last example can demonstrate the problem of
employing these rates in a simple model. Jassby and Goldman (1974) mea-
sured potential and actual growth of phytoplankton, and calculated mortality
by difference. They then estimated mortality due to sedimentation and to
grazing and found that the sum of these two processes was much less than the
calculated total mortality. They concluded that natural cell death is the main
cause of phytoplankton mortality. If they are correct, their analysis demands
a complete revision of the way we think about lakes. But is it right?
If we look more closely at their grazing calculation, we see that they took
data for the filtering rates of individual cladoceran and calanoid zooplankton
and summed these to get community grazing rate. This process ignores yet
another odd discrepancy in feeding studies. Haney (1971, 1973) developed a
technique to measure the grazing rate of zooplankton communities and indi-
vidual crustacean zooplankton in situ. He found that the measured total graz-
ing rate is much higher than the summed total of the individuals. Apparently,
rotifers, nauplii, protozoans, and other organisms ignored in the analysis of
Jassby and Goldman (1974) are responsible for much of the phytoplankton
mortality. We cannot leave inconvenient animals out of our analyses simply
because they are less studied and therefore presumed to be less important.
I could go on, but I think I have given enough examples to show that every
aspect of feeding behaviour of zooplankton is uncertain. I conclude that,
although the data are uncertain enough to encourage more study of zooplank-
ton feeding, they are not nearly good enough to be used in systems models.
Ecologists have developed a flourishing scientific sub-discipline to address
the feeding behaviour of zooplankton, and this work by and large treats the
animals as wholes, not as the sum of many mechanisms. However, that excel-
lent work cannot be combined reliably into reductionistic models of lake
ecosystems.
The Reality of Systems Analysis
Systems thinking now dominates ecology. It has convinced us quite
rightly that a reductionist theory must embrace all significant components of
113THE REALITY OF SYSTEMS ANALYSIS

the system and all significant interactions between these components. We
should also recognize the futility of trying to measure all those components
and interactions. Those who attempt to quantify them (as opposed to those
who build the interactions into a simulation model) will be overwhelmed by
the number of phenomena that need to be described quantitatively long before
the simplest ecosystem can be modelled.
Modellers have accepted this limitation and are usually content to fill the
gaps with interactions patterned after Michaelis-Menten kinetics, with arbi-
trary but reasonable constants, and empirical relations. Patten (1975) even
argues that we do not need good data for systems analysis, because the mod-
els work perfectly well without good data, so long as we refrain from testing
them against observation. However, if we wish to apply the model, we always
take short-cuts, replacing reductionistic descriptions with arbitrary rules or
empirical relations of at least some components. When we do so, we imply
that reductionism is inadequate.
One result of our recognition of these limits is the disorientation of exper-
imental ecologists. We do not have enough faith in the systems analysis mod-
els. On the other hand, we have lost our traditional, if naive, belief that there
is a key interaction and the ecologist fortunate enough to stumble on it would
produce the theory we seek. We have lost faith in our ability to generate use-
ful theories. Consequently, there is a large element of ‘going through the
motions’ in contemporary studies of ecological interaction.
This raises some interesting questions. Why do we do it? Why are zoo-
planktologists satisfied with bad estimates of every vital parameter? And
why are other ecologists similarly satisfied? I think the reason we expect
so little from science is that we have not yet realized that our job is to pre-
dict not to describe. We have not yet come face-to-face with our failure to
do effective ecological science. If we had effective, interesting, ecological
theories we could be testing them, and they would dictate the accuracy
required of our measurements. In the absence of theories, we can be as
careless as we like.
Conclusions
Now I think we can see what the computer has done for us. It has turned a
vague, semi-philosophical question of apparently little relevance to the work-
ing biologist into a practical, methodological problem. The computer gave
reductionists the tools required to approach an ecosystem as the sum of its
parts, and it allowed us all to discover that these tools are, and will always be,
inadequate. When ecologists realize what has taken place, they will take
114 REDUCTIONISM VERSUS HOLISM

renewed interest in the long-forgotten debate between holism and reduction-
ism, and this time the debate should have a different resolution.
There are two morals to my story. First, if ecology is to develop quickly
into a science that develops by testing its theories, we must recognize the
limitations of reductionism and put more effort into the holistic approach to
living systems. To make predictions about the future states of a system, we
must study the properties of that system, not those of its parts. Secondly,
philosophers and historians of science could play an important role in the
development of science. But to play this role, they must learn to communicate
with working scientists. They must find the right words with which to convey
their insights. This may mean that they must learn enough about day-to-day
science to convert their insights about the methodology of science into a
language scientists understand.
115CONCLUSIONS


VIII Sources of Ecological Creativity
“Science proceeds by revolution and not by
addition, pure and simple.”
Claude Bernard
[Le Cahier Rouge (1860)]
Philosophers of science have long distinguished between “the context of dis-
covery” of theory and “the context of justification”. The former, which is the
topic of this chapter, is concerned with the sources and modes of scientific
creativity. The latter deals with the evaluation of existing theories. Although
both phases are important to science, most writers treat questions about justi-
fication, rather than questions about the nature of scientific discovery.
Justification has received more attention for two reasons. First, it is the
public phase of science (Peters 1991a). Scientists are obliged to provide
scrupulously honest accounts of the tests of their hypotheses, but they need
not explain what inspired those tests or hypotheses. In fact, the rational devel-
opments of an hypothesis or test described in the introductions of scientific
papers are usually part of the context of justification and bear little relation to
the actual events surrounding the creation of the theory (Caws 1969). As a
result, the scientific literature does not lend itself to the study of creation.
Second, justification has received more attention because it has attracted and
yielded to the great philosophers of science, from David Hume to Karl
Popper. The same thinkers have placed far less emphasis on creativity, allow-
ing that discovery is a personal matter best left to the individual scientist and
his or her muse (Popper 1934).
The Challenge of Creativity
This uneven emphasis on creation and justification should be a matter of
some concern for working scientists. The evaluation and application of exist-
ing theories is relatively straightforward. We are by and large competent
technicians, distinguished by some modest originality, above average intelli-
gence, and a highly specialized education. This combination allows us to
function effectively in the context of justification. Our problem is more likely

to be the creation of clearly stated theories that are interesting enough to
warrant the effort of justification.
The problems of how to identify the hypotheses that will receive the
weight of our highly developed evaluative abilities should be addressed by
those philosophers who deal with the context of discovery, but that literature
is not very helpful in practice. Many of the writers who treat scientific cre-
ativity are visionaries who think that the object of their studies is too sublime
to be fully grasped (Caws 1969). As a result, their writing deals with concepts
that are too ill-defined for application. For example, we learn from Koestler
(1969) that there is a ripeness to ideas, but we are given no advice about how
we are to find these ideas or how we can compare maturities of ideas once
they are found. Koestler also tells us that we may expect a creative flash of
insight, but not how we can precipitate such a “Eureka act”. Most accounts of
scientific creativity give space to genius, imagination, and intuition (Morris
1966). The problem for working scientists is not that we mistrust the value of
these intangibles, but that an appeal to them is no help in our day-to-day work
(Caws 1969). Like most scientists, I recognize I have too little of these
virtues, and I gain nothing on being told I need more. What would be useful
are instructions about how to make my little store of imagination grow, or at
least how to use it effectively. The metaphors are simply unhelpful in that
regard.
Sociologists of science have approached the problem of creativity by plac-
ing science and scientists under the microscope and examining creation as a
phenomenon. They examine the lives of distinguished scientists to determine
if some similarity of education or environment might explain their stature.
From such studies, we learn that Nobel laureates tend to come from the mid-
dle class, studied with other Nobel laureates and worked at only a select few
universities (Zuckerman 1977). Francis Galton (1875), one of the first to
work on this topic, noted that distinguished scientists tended to be the eldest
child, had lost one parent before the age of ten, and had become strongly
attached to the other. Galton also emphasized the role of family and heredity
in genius. There is some indication that left-handedness may be more com-
mon among great scientists. Unfortunately, this information comes too late
for most of us. Our genetic dispositions, childhoods and formal educations
are already set in an unredeemable past.
There are some hints as to what we might do to be more creative. Caws
(1969), following Galton, suggests that creative scientists are critical of cur-
rent views in the science and suspicious of dogma. Koestler (1969) suggests
that we not resist new ideas, but try to prepare our minds to accept them. We
should consider alternatives (Chamberlin 1890, Platt 1964). The notable suc-
cess of scientists who switch fields suggests that periodic changes in research
118 SOURCES OF ECOLOGICAL CREATIVITY

topic are likely beneficial too. These steps are suggestive, but far from a cook-
book for creativity.
In considering this literature, I came to the conclusion that it has limited
meaning for most scientists because it deals primarily with scientists of
heroic dimensions: Galileo, Newton, Einstein, and Darwin. Even the sociolo-
gists of science limit themselves to such distinguished individuals that the rel-
evance of the conclusions for the rest of us must be suspect. For example,
Zuckerman (1977) deals with Nobel laureates, but figures in Price (1986)
show that only one in every one to ten thousand scientists can hope for such
achievement. What are the rest to do?
It can be argued that the research of the bulk of scientists simply does not
contribute to their science (Glaser 1964, Cole and Cole 1972). Statistics show
beyond the shadow of a doubt that the literature is dominated by a relatively
small number of scientific authors. These are the leaders who write most of
the papers, fill most of the best journals and receive most of the citations. At
the other extreme, there are a large number of authors who write but one or a
few papers, who are never cited, and who are relegated to lesser journals with
tiny readerships (Price 1986). The rest of us, mid-level career scientists,
could accept that we are unlikely to contribute creatively (Cole and Cole
1972) and resign ourselves to the important role of the audience that gives the
great scientists the recognition they have earned (Merton 1968). However,
most scientists legitimately hope to do more.
The Existing Literature as an Inspirational Device
Medawar (1967) suggested that scientists almost always draw their inspi-
ration from the literature of their science and their own scientific observa-
tions. Falling apples, swinging chandeliers, and slopping baths are therefore
as atypical as instances of scientific insight as Newton, Galileo, and Archi-
medes are atypical of scientists. Most of us do not write revolutionary papers
that redimension our sciences, and even scientific revolutionaries do so
rarely. Instead, most research is directed to solution of “puzzles” thrown into
relief by the paradigms of our sub-disciplines (Kuhn 1962, 1970).
Most of the remainder of this essay elaborates Medawar’s position using
the ecological literature. My concern is to provide models for scientific cre-
ativity that are more familiar and more functional for the majority of working
scientists, by providing more typical and more prosaic examples of the dis-
covery of pattern in nature. Although I have no authority to judge their appli-
cability beyond ecology, I would be surprised if scientific creativity in other
disciplines were remarkably different.
119THE EXISTING LITERATURE AS AN INSPIRATIONAL DEVICE

Dissection. The most common approach to the identification of relevant
problems in ecology has been the dissection of large things into manageable
parts. For example, the literature identifies many processes as important: the
phosphorus cycle in lakes, the productivity of forests, and the interaction
between plants and herbivores. Since these topics are too vast for a single
research project, we study some component of the total process that is inter-
esting and that we are competent to handle — plankton in a beaker, stomatal
gas exchange, or the feeding preferences of moose.
Fig. 27 provides a model of the rationalization for such studies (Rigler
1982a). The model provides a rationale for an almost limitless number of
small studies providing bits of previously unknown information about the
world around us. It can be used over and over again so as to help scientists
find new topics for research, and so continue playing the game. Science of
120 SOURCES OF ECOLOGICAL CREATIVITY
Fig. 27. The safe and easy game of ecology. Scientific research programs can be iden-
tified by choosing some population, species, community or ecosystem as X, and then
selecting the appropriate words and phrase in brackets. (From Rigler 1982a)
X is a very
abundant community
unusual type of species .
important ecosystem
species
Although similar communities have been studied,
ecosystems
as have some aspects of X
(references, to demonstrate knowledge of the literature) ,
feeding
production
X’s physiology has not.
Pb content
et cetera
Therefore I chose to study
one of the last set .

this sort is not particularly challenging, but it may be essential for normal
science.
Mechanism. Mechanism is a special case of dissection. Mechanistic anal-
ysis arranges other components of the system explicitly in flow diagrams and
simulation models, or implicitly as a mental construct in the minds of the pro-
ject directors. The object of study is important only because it is an essential
part of the whole. This device was effective in realizing the aims of NASA
and the Manhattan project, where a final, tangible goal was well recognized
and the parts were developed for this application.
Mechanistic analysis has also produced a number of fine descriptions of
various ecosystems (e.g. as part of the International Biological Program). It
seems to have been less successful in this area, perhaps because the problems
the ecological teams were to confront were less clear than bombing the
enemy or walking on the moon.
Although this essay is most concerned with indicating some of the ways
that working ecologists find their problems, the preceding chapter obliges me
to point out a difficulty with mechanism. The rationale assumes that the
importance of the process does not lie in the isolated parts, but in the whole.
If the program is to succeed, someone must eventually put the disconnected
parts together. This synthesis has proven remarkably difficult (Brown 1981).
Dichotomies and categories. Dissection and mechanism are reductionis-
tic approaches to theory building in ecology, but there are holistic alterna-
tives. One of these involves a thorough classification of the phenomena and
subsequent hypothesis about the characteristics of the classes. For example,
Robert MacArthur repeatedly identified extreme behaviours and then used
these extremes as dichotomous categories for the classification of less
extreme behaviour of the same type (Schoener 1972, Kingsland 1985). Thus
we have opportunist and equilibrium species, generalist and specialists, and
coarse- and fine-grained environments. A similar approach takes the
extremes as the poles of an axis, as in the r–K continuum in life history
theory. In all cases, the approach involves two steps: first an identification of
the defining categories or axis, then an examination or comparison of the
different characteristics associated with the resulting categories or ranks.
There is no reason to limit this approach to dichotomous categorizations.
Southwood (1988) advocates a trichotomous division of the habitat in terms
of disturbance, adversity, and biological interaction. MacArthur (1972)
believed that future developments in ecology would depend on a two- or
three-way categorization of the habitat and its potential inhabitants such that
ecological theory would take the form: in environments of type B, organisms
of type X will have characteristics P, Q, and R. The biome approach goes
further still.
121THE EXISTING LITERATURE AS AN INSPIRATIONAL DEVICE

Analysis of variance. Categorization is often qualitative, but an analo-
gous quantitative approach is available. This uses the power of common
statistical procedures to identify tentative patterns which can then be evalu-
ated by further tests. Again one selects a significant topic from the literature,
but now interest is limited to quantitative estimates of the process of interest.
For example, primary production may represent the productivity of lakes and
fish catch that of the sea. One then collects as many estimates as possible and
estimates the variance. The variance quantifies the extent of our ignorance
and is addressed by determining what other factors explain a significant frac-
tion of this variation. Qualitative factors are assessed by expressing these
variables as simple categorical variables and applying analysis of variance
(ANOVA) to determine the explanatory power of the categorization. The
effects of continuous variables can be estimated by regression techniques,
which are themselves minor variants of ANOVA, and mixed models with both
categorical and continuous variables can be developed with analysis of
covariance (ANCOVA). Downing (1991) found that 50 % of the statistical
comparisons between ecosystems used one of these variants; a further 40 %
relied on visual inspections of graphs or tables, which are simply the non-
statistical analogues of ANOVA. This empirical approach has been particu-
larly effective in developing nutrient response models in limnology (Peters
1986) and allometric models in autecology (Peters 1983, Calder 1984).
Extensions, additions and modifications. Almost all relations in ecology
are unsatisfying because the uncertainty of prediction is too large, because the
domain of the relation is too small, or because the original work suffers some
potential bias or other shortcoming. These limitations have long been fruitful
areas for new research. For example, the phosphorus-chlorophyll relation
first proposed by Sakamoto (1966) was improved by Dillon and Rigler
(1974a) who expressed the relation statistically, showed that it applied out-
side Japan, added more lakes to the data base, and, following Vollenweider
(1968), related it to the problem of phosphorus abatement. Since then, more
than 60 relations (Peters 1986) have been added to show that the relation
holds or to modify it for use under a variety of conditions or regions. Such
modifications suggest different mathematical forms for the relation
(Strasˇkraba 1980, McCauley et al. 1989, Prairie et al. 1989), different param-
eters (OECD 1982), different measures of algal biomass (Nicholls and Dillon
1978), and additional variables (Smith 1982, Canfield et al. 1984). Some
authors have concentrated on developing the largest possible data set (Can-
field and Bachman 1981), others on explaining the variation in the existing
sets (Carpenter et al. 1985).
Similar developments have occurred in allometry. Beginning with Rub-
ner’s study of respiration in dogs (Kleiber 1961), there has been a consistent
122 SOURCES OF ECOLOGICAL CREATIVITY

tendency to increase the range of weights, the number of species, and the
number of higher taxa represented (e.g. Benedict 1938, Brody 1945, Hem-
mingsen 1960, Pagel and Harvey 1988). Robinson et al. (1983) considered
temperature as well as size; McNab (1980) has suggested that life styles and
food habits be considered too. Some few champion other models (Economos
1979, Smith 1980, Seim and Saether 1983) and many have argued for differ-
ent statistical treatments (Zar 1968, Jolicouer and Heusner 1971, Harvey and
Mace 1982, Ricker 1984, Pagel and Harvey 1988). Moreover, successful
models, like the phosphorus-chlorophyll models and size-metabolism
models, have encouraged the development of similar models predicting other
responses from the same independent variables or from correlates of these
variables (Peters 1983, 1986).
The amazing fertility of extension, addition and modification is not lim-
ited to large scale regression analyses. Indeed, limnologists have so long
insisted that each lake and each species is different, that we can safely claim
to be doing science simply by repeating earlier research in a previously unin-
vestigated region, or with a slightly different organism, or at a different time
of year. Extensions to new domains, additions of new variables, and modifi-
cations to existing analyses and models are all implicit elements in the model
for normal research in ecology in Fig. 27.
Technologies. The application of a new technology to the traditional prob-
lems of aquatic ecology is another way to generate a research problem in
ecology. This allows us to profit from advances in our sister sciences includ-
ing the traditional areas of electronics, analytical chemistry, mathematics, and
statistics, but also molecular biology, bioengineering, and computer science.
For example, we can now work at higher levels of sensitivity in the analysis
of the contaminants of our waters, we can identify the genetic structure of our
study populations, and we can collect and analyze data at a rate which was
unimaginable only a few years ago. We can only expect these fruitful inter-
actions to grow. Those ecologists who are technologically competent can
therefore look forward to long, profitable, and potentially much cited careers
dedicated to the translation of advances outside of ecology to ecological
applications.
Complications. Still another mode of creation in ecology has involved the
further complication of existing theoretical models by allowing greater com-
plexity. The history of the logistic curve and its descendants provides an
example (Hutchinson 1978, May 1981). The logistic is itself a modification
of the equation for exponential growth over time (t) of a population contain-
ing N individuals, where r is the growth rate constant:
δN/δt = rN (9)
123THE EXISTING LITERATURE AS AN INSPIRATIONAL DEVICE

The logistic sets an upper limit to this population by assuming that population
growth may be described as sigmoid growth to an asymptote (K) so that the
rate of increase slows as the asymptote is approached:
δN/δt = rN(1 – N/K) (10)
When it became apparent that few populations grew according to this for-
mula, the formula was complicated by the addition of other terms to include
other factors. Thus, the basic equation could be modified (Hutchinson 1978)
to accommodate the lag whereby population growth at time t is a response to
population size at time t –1:
δN/δt = rN(1 – N
t–1
/K) (11)
Alternatively, one could presume that the asymptote varied in time and
thereby replace K with a function of time, or a function of resource supply.
This was developed further by Lotka and Volterra to treat the effects of a
predator population (containing P individuals):
δN/δt = Nr(1 – N/K) – α NP (12)
δP/δt = –Pd + β NP (13)
where α NP describes the decline in prey growth as a result of predator-prey
encounters. The growth of the predator population is instead assumed to
represent a balance between the predator death rate, d, and some positive
function of predator-prey encounters, β NP. These equations can be elabo-
rated much further (May 1981).
A further set of equations based on this model were developed by Volterra
to describe population growth of the ith species as a function of both its own
population size and the effect of 1 to j competing species, each characterized
by a competition coefficient α
ij
:
δN
i
/δt = r
i
N
i
(1 – N
i
/K
i
– ∑
j
α
ij
N
j
/K
j
) (14)
This led to the very rich conceptual developments of MacArthur, Levins, May
and others, treating communities as matrices of mutual competition coeffi-
cients.
Since an infinite number of equations could replace the simple relations of
Verhulst, Lotka and Volterra, these developments could go on indefinitely.
Moreover, since there are also an infinite series of alternatives to these simple
models, we could expect to see creative programmes in ecology eventually
developing parallel to the logistic mainstream.
Not all complications need be so mathematical. Some can be largely
qualitative. For example, models of zooplankton filtering rate (Chapter VII)
124 SOURCES OF ECOLOGICAL CREATIVITY

began with the assumption that the animals were simple pumps and filters.
Gradually researchers have uncovered a far greater range of complications
showing that the feeding rate is sensitive to temperature, pH, oxygen, food
particle concentration, particle size, particle type, experimental methodology,
animal feeding history, age, size and so on (Peters 1984, Lampert 1987). Sim-
ilar developments could be sketched for any physiological process. Another
limnological example, but one which invokes community ecology, is pro-
vided by the size efficiency hypothesis (Hall et al. 1976, Peters 1991b, 1992)
which began as a rather straightforward case of predator-mediated competi-
tion between large and small animals (Brooks and Dodson 1965), but which
has subsequently been developed, extended, and complicated by considering
the effects of invertebrate predators and their vertebrate predators (Dodson
1970), of food particle size, of starvation resistance in juveniles (Tessier and
Goulden 1987), of interference due to larger particles, and of differential
capacities to avoid entanglement (Webster and Peters 1978, Gliwicz and
Siedlar 1980, Porter and McDonough 1984). The paradigm has built on the
original broader base of Hrbácˇek et al. (1961) to include a host of top-down
effects (McQueen et al. 1986) and may be considered a forerunner of bio-
manipulation (Shapiro and Wright 1984). Ecologists thrive on complication
and therefore an easy step to creativity is to develop still another flourish for
the existing models.
The Danger of Conventionalism
It is not my intention to describe all the ways that ecologists come to their
new ideas. Instead of developing this list further, I want to end by considering
a problem that results if these are the only models for creative developments
in ecology and to point to a potential solution to that difficulty.
The flaw is that these developments are all based on the overriding
paradigms and dogmas of contemporary ecology. If we use any of these mod-
els we further entrench prevailing ideas. Therefore before we continue any
line of development we should invoke our critical capacities to determine if
that line merits development. In some cases, including limnological models
of lake nutrient-response models and autecological models based on allome-
try, I think a fair evaluation would encourage such development. These are
good theories with considerable promise for growth and application. In other
cases, the logistic and competition theory being two, conventional creativity
has led to many publications and citations, but not to predictive power or the
resolution of problems that such power allows. I suspect that the normal cre-
ative modes of ecology sometimes simply extend the life of constructs which
125THE DANGER OF CONVENTIONALISM

have not borne fruit in the past (Brown 1981) and are unlikely to do so in the
future.
The traditional questions and topics of our science can be fruitful sources
of inspiration for new research. They can also be intellectual traps. We risk
entrapment when we allow the science to become so self-defined that the only
interesting questions are those which we have always asked, but failed to
resolve. This is the final retreat to scholasticism, it is part of what Pramer
(1985) called “terminal science”, and it is the opposite to Medawar’s (1967)
conception of science as “the art of the soluble”.
Creative Alternatives for Normal Ecology
If we are to escape from the trap of terminal science, we need a means of
introducing new questions. At first, this may seem to require a higher level of
creativity than the techniques mentioned above, but there are some ways of
finding new questions which need not lie outside normal science.
Syllogisms and analogies. One spectacularly successful technique to
encourage creativity is that of Hutchinson (1978; see Kingsland 1985 for
discussion). This consists in the identification of a syllogism — that is of
some logical, usually algebraic or graphical, model — from some area out-
side of ecology, and application of this syllogism to a problem in ecology
which is hypothesized to be analogous. This approach appears in analogies of
ecology with economics, game theory, physics and genetics. It is frequently
used in competition theory, optimality theory, and evolutionary theory, and
has been characteristic of the powerful school of ecological thought domi-
nated by Hutchinson, MacArthur, and their intellectual descendants (Fretwell
1975, Brown 1981). It is especially useful for those who have interests and
abilities which extend beyond ecology.
The process has two relatively independent phases. The logical implica-
tions of the model are elaborated in a deductive phase. Since this elaboration
is a purely logical question, this phase requires no empirical input and there-
fore this is a major activity for theoretical ecologists. Once the deductive
phase is well advanced, the research enters a hypothetical phase in which the
model is hypothesized to apply to some ecological phenomenon. Identifying
such a phenomenon may prove difficult or impossible, and some of these log-
ical models leave the theoretician with a solution in search of a problem, like
the logistic and its descendants.
Because this approach is based on analogy and metaphor (Oster 1981), it
is only as good as the degree to which the analogy holds. In practice, the anal-
ogy is rarely perfect and considerable ingenuity may be required to fit some
126 SOURCES OF ECOLOGICAL CREATIVITY

ecological observations to the logical model (Maynard Smith 1972). How-
ever, even if the model cannot be applied to nature, its logic may become a
part of theoretical ecology. For example, this seems to have happened to the
theory of limiting similarity, whose terms cannot be applied to nature (Brown
1981) but which remains a favourite point of discussion for theoretical ecol-
ogy (Abrams 1983). Sometimes, as in optimal foraging theory (Beatty 1980,
Stephens and Krebs 1986, Gray 1987), one may be in a position to decide
which model applies to a particular case only after making the observations
that the models purport to predict. Such models only tell us what we already
know.
The syllogism has proven a useful prod to ecological creativity, but it has
also introduced a series of logical arguments into ecology which are either
impossible to apply to nature, or can only be applied after the fact. These are
the tautologies that confound ecological texts, courses and journals, and slow
the development of a powerful science (Peters 1976, 1991a).
A return to application. There is another way to stir the creative juices of
ecologists and thereby help them to isolate appropriate research problems.
This also involves material from outside the traditions of the science, but it
has been less widely recognized than syllogism and analogy.
I suggest that we step back from our library desks and lab-benches, that we
put aside our field notes and learned journals, and instead take a long look at
our lakes, our seas, our forests and our fields. We should ask ourselves what
are the most obvious, first questions that we would put to such a system. We
might wonder, for example, if the water in a lake is good to drink or if it is
warm enough for swimming. We might wonder what fish live there, where in
the lake they are found, how many we can catch, and if they are safe to eat.
We may have other concerns like how much water we can drain from the lake
for agriculture or how the fish will respond to water level controls, industrial
development, and acid rain. There are a host of questions like this, questions
we once asked, but of which we lost track in the course of the professional-
ization and professorization of the science.
Often, we have turned our backs on these obvious questions to ask ques-
tions like “How many eggs does a copepod carry?”, ”What is the successional
sequence of algal species in the lake?”, “What are the names of the animals
which the fish have in their stomachs?”, “How fast is radioactive phosphate
absorbed by the plankton?”, and “How much does a water flea excrete?”.
These may be important questions. We may need to know these things to
answer the obvious questions. Whether this is so or not, we have lost the con-
nection between these obvious questions that first attracted many scientists to
ecology and many contemporary ecologists to their specialities, and those
which our scientific training has subsequently encouraged us to pose.
127CREATIVE ALTERNATIVES FOR NORMAL ECOLOGY

The benefits of practice are widely recognized. De Solla Price (1986)
made the point in a general context in his last paper, On Sealing Wax and
String. Box (1976) argued that the interchange of theory and practice was an
essential element in the genius of R. A. Fisher. Vollenweider (1968) showed
that it is possible to contribute powerfully to limnology, while ignoring the
traditions of the science. Application is more scientifically valuable, more
socially essential, and more intellectually challenging (Rigler 1975a, 1982a,
b) than most of normal science.
Therefore my last suggestion to increase our creativity is a simple one. It
is to return to the questions of the layman, or better to the questions of a child.
In doing so, we may be able to see our subjects with fresh eyes, lighting again
the creativity that effective science demands, and simultaneously helping
confront the ever-growing problems of humanity.
128 SOURCES OF ECOLOGICAL CREATIVITY

IX Empirical Limnology
“No scientist is admired for failing to solve problems
that lie beyond his competence. The most he can
hope for is the kindly contempt earned by Utopian
politicians. If politics is the art of the possible,
research is surely the art of the soluble. Both are
immensely practical-minded affairs.”
Sir Peter Medawar
[The Art of the Soluble (1967)]
In previous chapters, I defined science and theory to emphasize the impor-
tance of theory in telling us what the world is like. According to Popper, sci-
ence does this by telling us what the world is not, so that we can discount
logically possible observations that the theory tells us are unlikely to occur. I
then detailed my growing unease as I searched for such theories in ecology
and instead discovered that many ecological constructs were not very infor-
mative in this sense. I looked for guidance to my speciality and to the general
field, but was again disappointed. They did not seem to measure up, either to
Popper’s criterion of demarcation or to the growing environmental challenge.
Initially, my disillusionment was profoundly disquieting, but eventually it led
me to resolve the issue by putting aside the traditional topics of my research
to focus on socially relevant, holistic, empirical studies of lakes. That change
in approach is the topic of this chapter.
Social Demands and Scientific Supply
For many years, eminent ecologists have warned that the present course of
society will lead to certain disaster for civilization. For the most part, society
has ignored them.
I was never an eco-preacher — I am much too selfish about research
time — but generally I support that mission and I have wondered why the
excellent arguments of our leading ecologists were ignored. I resolved the
issue, at least in my own mind, by recognizing that the questions ecologists
were answering differed from those society was asking. The eco-preachers

made predictions about the fate of the biosphere, but society was asking ques-
tions about populations and ecosystems. Ecologists would eagerly argue for
global restructuring of the economy, culture and politics, but were often mute
in discussions about cleaning a river or managing a fish stock. The ecologists
argued, with some justification, that biospheric questions were more impor-
tant. Society wondered, equally legitimately, why it should accept ecologists’
solutions to big questions, when it was apparent that ecologists could not
resolve local issues.
This ecological credibility gap grew when society realized that most ecol-
ogists had not been trained as biospheric experts. Ecologists trained to
address much smaller questions, yet, when asked to make a specific predic-
tion about a particular population or ecosystem, we could rarely do it. Even
more rarely did we agree with one another.
The standard reply to a request for advice was in effect, “Give me
$100 000 per year for twenty years to study the issue and I’ll tell you what
happened.” Society caught onto this trick pretty quickly. Instead of asking
one ecologist, it asked a suite of n ecologists, and out of the n different
guesses that resulted, it picked the guess that was most economically or polit-
ically expedient. This tactic simply obfuscated the process of finding scien-
tific solutions to societal problems, delaying science by confusing both
researchers and society.
My discovery that ecologists’ answers were mismatched with society’s
questions led me to a personal choice to start predicting what society wanted
to know. My decision to change research directions after many years was
hard, but I found it even harder to develop useful theories. I could, with mod-
est intellectual effort and considerable hand-waving, invent a new theory, but
I could not guarantee its success in application. I therefore focused on finding
the approach most likely to create a “useful” theory. In this context, “useful”
means predicting what society wanted to know.
Pessimists and Optimists
There seemed to be two distinctly different methods of attacking a scien-
tific problem, which I shall call, for the present, the way of the optimist and
the way of the pessimist. They can be distinguished by the way they address
difficult scientific problems. The optimist says “I am going to solve it.” The
pessimist says “It is not soluble in its entirety, therefore I will solve a little bit
of it.”
Imagine that each school wanted to predict when an alarm clock would go
off so that they might control this device to their advantage. The optimist
130 EMPIRICAL LIMNOLOGY

would study the behaviour of the clock under a variety of conditions and look
for regularities or patterns. The pessimist, believing that the clock could
never be really understood that way, would instead offer to take the clock
apart: “I will study each screw, and each cog wheel. I will take apart the cogs
and see how they are made, I will study the shape of the teeth, and the
scratches on the casing; I will look at the structure of the brass, steel and lead.
Then I will begin to study the interactions between and among these parts,
and slowly, painstakingly, I will come to understand the clock.” Given the
information at hand, we have no way of knowing which approach is better.
The scenario supposed that we could not predict the clock’s behaviour, and as
long as this is so, we cannot judge the success of the different approaches.
Any choice between them is whimsical, unless we can appeal to a broader
experience. We need a theory of scientific development.
Testing the alternatives. Luckily for me, I once taught a course on the
history of biology, and I was able to cast my mind over examples of the
invention of past theories. To my delight, I discovered that they all came
about in the same way.
If we were to look over the history of branches of science that have
achieved successful theories, we would discover a remarkably consistent
story. It runs as follows:
(1) identify the system about which you want to make predictions;
(2) observe the behaviour of that system and look for pattern;
(3) express the pattern rigorously and, if possible, quantitatively to yield
an empirical theory;
(4) try to explain why the theory works, with an analytical or explanatory
theory.
Thus Copernicus built on patterns that had been observed by astronomers
from Babylonia to Tycho Brahe, and Newton “only” completed the quantita-
tive description of those patterns begun by Kepler and Galileo. That arche-
typical scientific advance did not reach the status of explanation until a new
generation saw it as philosophically true (Chapter III). Dalton saw that chem-
icals combined in fixed ratios, Mendel tended his peas, and Darwin visited
the Galapagos. Even the most elegant analytical theories began as humble
empirical patterns.
This is not to say that the discoverers of pattern had no preconceptions or
working hypotheses. They undoubtedly did. Nevertheless, the initial work
was observational and the first achievement was the identification of patterns
in those observations. Consequently, most of the creative approaches of nor-
mal ecology (Chapter VIII) aim to detect pattern. That is an appropriate focus
for a young science.
131PESSIMISTS AND OPTIMISTS

One of our problems in ecology is that many ecologists have decided that
this standard scientific strategy is inappropriate for ecological systems. Ecol-
ogists have placed their trust in alternatives. Perhaps the most popular has
been reductionism or mechanism, whereby we dissect a system we see as too
complex and study the parts in isolation. An alternative technique, less used
but still much honoured, is the “a priori road” (Forbes 1887) whereby we
intuit bold hypotheses for subsequent test. Still another technique creates
elaborate intellectual universes and ecosystems of concept or algebra and
then seeks parallels between those intellectualisms and the real world. These
approaches are popular and attractive, but the weakness of ecological theory
in the face of the environmental crisis argues tellingly against them.
Holists and reductionists. I have made two important points. Successful
investigators did not begin by dissecting the system of interest; and the origi-
nal theories did not pretend to convey understanding. In other words, histori-
cal evidence suggests that the way to construct theories has been holistic and
empirical. There is really only one way to create new theory and that is the
way of the optimists.
My perception that there were two ways to create theory was based on a
misapprehension. The source of this misapprehension was also revealed by
our collective experience in building ecological theories. After a pattern had
been identified empirically, other investigators were inspired to explain why
this holistic theory worked. They might tear the system apart and relate the
behaviour of the whole to the behaviour of the parts. They might attempt to
rephrase the theory in words that conveyed understanding, and if successful,
they might gain far more credit than the empiricist who first described the
pattern. The pessimists then were not trying to build a new theory or to offer
useful new predictions. They were trying to explain theories that already
existed. Like many ecologists, I had confused the two processes and now see
my early years in research as a misapplication of the pessimist’s piecemeal
approach to the creation of new theory, rather than to the explanation of exist-
ing theory.
In more conventional terms, the ecological optimists are called “holists”
and the pessimists “reductionists”. In general, scientists are much happier
with a reductionist, explanatory theory than with an empirical, holistic theory,
even though both may be equally predictive. As a result, most researchers
approach their material as reductionists. However, since I was concerned with
producing predictions that contemporary ecological theory could not give,
there was no reason for me to work like a reductionist. My decision was there-
fore to reject the reductionist approach and to become a holist and an empiri-
cist. Thus after twenty years, I stopped being a scientific pessimist and
became an optimist.
132 EMPIRICAL LIMNOLOGY

What to Predict?
Once I had decided to hunt for a theory, I had to decide what the theory
was going to be about. What system would I study and which of its proper-
ties would I predict? The system would clearly have to be a lake, because I
had no expertise about anything else. At the time, there were a number of
questions people asked about lakes: Is it green? Does it smell? Are there fish
in it?
At first, I balked at the idea of addressing topical issues in society. The
professor in me, the old scientist, grey, wrinkled, and paunchy, wanted to
say “No, you must stick to pure science because only basic research yields
new theories.” The concerned scientist, interested in society’s problems and
society’s image of ecologists, countered by asking how I could hope to close
the credibility gap if I insisted on making predictions that were irrelevant to
society.
Luckily, history again came to the rescue. The old professor was wrong.
Many contributions to scientific theory come from attempts to answer soci-
ety’s questions. Astronomical theory arose from our perceived need for a
calendar. The earliest chemical theory arose from our desire for eternal life
and for the ability to convert base metals to gold. More recently, the germ
theory of disease and our theory of spontaneous generation arose because
Louis Pasteur was hired to find ways to cure diseases of grapes and wine.
Claude Bernard, motivated by a desire to cure diabetes, developed our
theory of homeostatic internal regulation. The best examples of useful theo-
ries in ecology that I knew were models developed to predict commercial
fish catch by D. S. Rawson (1955) and later by Dick Ryder (1965, 1982).
Thus we have plenty of evidence to show that new theories can come from
applied problems.
The problem of which property to predict remained. Society wants pre-
dictions about many properties of lakes: taste, toxicity, algal abundance,
oxygen levels, contaminant fates, fish harvest, and so on. How does one
select amongst these properties? An answer to this question was given most
succinctly by the Nobel laureate P. B. Medawar (1967) in a review of Arthur
Koestler’s book, The Act of Creation. Medawar answered Koestler’s ac-
cusation that scientists ignore the important questions by saying that these
questions are too difficult. Successful researchers deal with soluble prob-
lems. Here was the answer I was looking for. I should try to make predic-
tions about the easiest problem I could identify. For a variety of reasons,
most of which concern personal competence and logistic support, the most
tractable problem of societal interest was the problem of predicting algal
abundance.
133WHAT TO PREDICT?

A Research Program in Holistic Empirical Ecology
How green is my lake? I had decided that we needed predictions. The his-
tory of biology showed that the holistic, empirical approach was most likely
to produce the requisite theory. Algal abundance looked easy, so that was my
first target and I did not mind in the least that society was interested in that
result. My approach would be to search for repetitive patterns in the algal
abundance of lakes and then to correlate these patterns to other lake proper-
ties, so that in future we would be able to predict algal abundance from those
properties.
The twin perils of complexity and reality. At this point, I had to be careful
not to fall into a dangerous trap. I must not allow myself to be distracted by
all the fascinating complexities of the system. For example, it was tempting to
start by counting all the species and races of algae in a series of lakes; or I
might have instead begun to speculate about the relation between the property
I wanted to predict and other equally unpredictable properties, like the rela-
tion between zooplankton and algae. These questions, and many others, are
quite capable of engaging armies of limnologists for generations.
To avoid such traps, I very quickly decided that the measure of algal abun-
dance for my purposes was the concentration of chlorophyll a. This choice
seems dangerously simplistic to the modern limnologist who knows that dif-
ferent algal divisions contain different pigments, and that the chlorophyll:
biomass ratio can vary enormously within a single algal species (Nicholls and
Dillon 1978). Indeed the choice is even more simplistic because we are not
even measuring chlorophyll a. All that I was trying to predict was the absorb-
tion of a particular wavelength of light by the pigments extracted from the
particles collected in a sample of lake-water.
The decision to study chlorophyll extracts put me in danger from another
trap, that I might become stalled in a lengthy consideration of the reality of
my measurements, just as I had once studied the reality of phosphorus frac-
tions. I escaped both pitfalls by applying a holistic, empirical approach. The
holist avoids dissection of the problem (i.e. the prediction of total algal abun-
dance) into its components (e.g. the prediction of algal species abundance).
The empiricist focuses on measurable properties. The amount of green colour
to be extracted from lake-water by a rigorously defined method is a measur-
able property of the lake. To keep my mind focused on the right approach, it
would probably have been useful to say that I was trying to predict greenness
rather than algal abundance. Not surprisingly, that was what society wanted
to know because that is what people see.
Success — the phosphorus-chlorophyll relation. I was now ready to look
at the data and find patterns in the greenness of lakes. Whenever one starts
134 EMPIRICAL LIMNOLOGY

such a search for pattern in nature, the obvious place to begin is the published
literature. When I did so, I discovered the problem was even more easily
solved than I hoped, because much of the work was already in the literature
waiting to be used.
In a study of 40 Japanese lakes, Sakamoto (1966) showed that, where the
ratio of total nitrogen concentration to total phosphorus concentration was at
least 12:1, chlorophyll concentration is closely correlated with total phospho-
rus. I immediately began looking for other lakes to test this relationship. I
found work by Deevey (1940) in Connecticut, by Edmondson (1969, 1972) in
Lake Washington, and by others elsewhere in North America. All of them fit
very well on Sakamoto’s plot. Peter Dillon (1973) confirmed the applicability
of this relationship in a further series in Ontario, and Wolf Scheider (1978)
did the same in a series of much smaller lakes. All the data fit the same rela-
tionship (Fig. 28).
Twenty years ago, the phosphorus-chlorophyll relation was a revolution-
ary discovery, so counter-intuitive (Elster 1958) that for a time it seemed
unpublishable. Since then, the basic positive response has been confirmed so
many times that the relation is taken for granted. Some even say that the rela-
tion is tautologically true, although I have never seen the deductive proof that
claim implies. Instead, I think the imputation of tautology indicates that the
135A RESEARCH PROGRAM IN HOLISTIC EMPIRICAL ECOLOGY
Fig. 28. The phosphorus-chlorophyll relationship in lakes. (From Dillon and Rigler
1974a)

relation is now considered self-evident. The phosphorus-chlorophyll relation
has become philosophically true, because it now is what we expect.
Phosphorus concentration. At first, we misinterpreted the data in Fig. 28
as a correlation between total phosphorus concentration at spring overturn
and mean summer chlorophyll, but now we would see this as a relation based
on seasonal mean of the phosphorus concentration. In either case, this is not
a particularly useful relationship because it is as much work to measure phos-
phorus as it is to measure chlorophyll and because there is no indication in the
correlation that we could manipulate chlorophyll by manipulating phospho-
rus. However, if we could predict phosphorus concentration from some other
easily measured property we might be getting somewhere; and if we could
relate mean phosphorus concentration to the quantity of phosphorus entering
a lake or something similar, it would be more useful still.
Vollenweider’s basic model. In 1969, Richard Vollenweider published a
model for substance budgets in lakes that purported to predict the concentra-
tion of any substance in lake water. The model is extremely simple. It treats
lakes as if they are open systems, each consisting of a single compartment in
steady state (Fig. 29). For the present discussion, the model can be specified
to predict average lake phosphorus concentration ([TP], in mg m
–3
):
[TP] = L/(q
s
+ σ z
m
) (15)
where L (mg m
–2
yr
–1
) represents the annual load of phosphorus to the lake’s
surface, and the other terms may be thought of as correction factors for other
processes. Mean depth (z
m
, in m) corrects for the depth over which the load
must be distributed; the term for areal hydrological load of water to the lake’s
surface (q
s
, in m
3
m
–2
yr
–1
or m yr
–1
) corrects for loss of phosphorus through
the outlet, and the sedimentation coefficient (σ, yr
–1
) corrects for the sinking
of phosphorus from the lake water to the bottom. If it worked, this model
could be combined with the phosphorus-chlorophyll regression (Fig. 28) to
predict the effects of phosphorus abatement and enrichment on the greenness
of lakes.
Modifications and reformulations. Vollenweider’s model appeared to
work very well for total phosphorus in relatively unproductive lakes, but not
in highly eutrophic lakes. However, the model had a more important flaw. It
required an estimated sedimentation coefficient to describe the net rate of
phosphorus sinking, and this constant is very difficult to measure directly.
Our solution was to reformulate the model slightly, replacing the hard-to-
measure sedimentation coefficient with a “retention coefficient” so that all
terms were measurable (Dillon and Rigler 1974b). If the total amount of
phosphorus entering the lake from all sources is J (the phosphorus load in
mg yr
–1
), and the total amount of phosphorus leaving the lake via the outflow
136 EMPIRICAL LIMNOLOGY

is J
out
(also in mg yr
–1
), the retention coefficient (R) is defined as the frac-
tional loss
R = (J – J
out
)/J (16)
R is therefore the net fraction of the incoming phosphorus which is apparently
lost to the sediments.
Given R, J and an estimate of the net amount of water flowing into the lake
from all sources (the hydrological load, Q, in m
3
yr
–1
), one can predict [TP]
in the lake water as
[TP] = (J/Q)(1 – R) (17)
Eq. (17) simply states that lake phosphorus concentration is equal to the
volume-weighted mean phosphorus concentration of all sources of water and
phosphorus (J/Q), corrected for net loss of phosphorus (1 – R). Dillon (1973,
Dillon and Rigler 1974b) then showed that this formulation could effectively
predict phosphorus concentration in lakes. These results were subsequently
confirmed in a set of smaller lakes by Scheider (1978).
137A RESEARCH PROGRAM IN HOLISTIC EMPIRICAL ECOLOGY
Fig. 29. The lake interpreted as a simple continuously stirred mixed reactor in which the
internal concentration at steady state is the result of a single source, the inlet, and
two sinks, the outlet and the sediments. (From Reckhow and Chapra 1983)

Predicting the components of the model. Unfortunately, the model de-
scribed by Eq. (17) is useless for many practical purposes. Because estimates
of J, Q and J
out
require vast field programmes, they are far more expensive
and time consuming than measurements of [TP]. An estimate of Q requires
estimates of the sum of the annual flows of all the i tributaries to the lake (Q
t1
,
Q
t2
, ..., Q
ti
), the sum of the water inputs from each of j “precipitation events”
(Q
p1
, Q
p2
, ..., Q
pj
), less evaporation (E) from the lake surface:
Q = ∑Q
ti
+ ∑Q
pj
– E (18)
An estimate of J requires estimates of the total phosphorus concentration
associated with these various inputs from tributary streams ([TP
ti
]) and pre-
cipitation including dry fallout [TP
aj
]:
J = ∑Q
ti
[TP
ti
] + ∑Q
pi
[TP
aj
] (19)
And estimates of R require all this plus an estimate of [TP] and water flow
in the outflow. Obviously, the approach would be much more useful if we
had a way to predict these components of the phosphorus mass balance. If
this were possible, we might be able to predict how green a lake would be
with minimal field work, perhaps without even visiting the lake. This be-
came our goal (Dillon and Rigler 1975).
Q was the easiest of all the components of the budget to estimate. Because
the quantity of water in streams is important for many purposes, hydrologists
had been measuring water flows for years and government sources were able
to provide maps describing the average annual flows of water per unit area of
drainage basin for all of Canada. The same sources provide estimates of evap-
oration from and precipitation onto lake surfaces. Analogous agencies can
provide the same information for other jurisdictions. We simply had to
assume that future flows would resemble past ones and we were able to pre-
dict the components of Eq. (18).
Predicting the phosphorus concentrations associated with this water was
more difficult, because that characteristic had been measured less frequently.
However, Dillon and Kirchner (1975) were able to collect enough estimates
from the literature to provide average figures for the amount of phosphorus
exported per unit area of watershed, given contrasting patterns of land use
and geology (Table 12). The values were only averages, the scatter is high,
and some land uses are not addressed, but the data were then the best avail-
able. As this approach eventually proved powerful, subsequent authors
(Reckhow and Simpson 1980, Prairie and Kalff 1986) have improved and
extended it.
Because P load is calculated as the product of hydrological load and phos-
phorus concentration, Eq. (19) requires estimates of the phosphorus in pre-
138 EMPIRICAL LIMNOLOGY

cipitation and dry fall (Q
p
[TP
a
]). Dillon (1973) estimated an average value for
this “aeolian input” (77 mg m
–2
yr
–1
), and we were surprised to find that in
many of our lakes rain, snow and dust could be the major source of nutrient
(Rigler 1974). Because there were many opportunities for error in our esti-
mates of aeolian input, Renata Gomolka (1975) took a much more careful
look at the phosphorus associated with rain and dust. Her work showed that
much of the phosphorus collected by shore-based funnel traps overestimated
the amount that falls on the lake surface. Nevertheless, her estimate (37 mg P
m
–2
yr
–1
) still identified the air as the major source of phosphorus for many
undeveloped lakes with small catchments. This still seemed counter-intuitive,
but subsequent work has confirmed both the magnitude of this input and its
biological reactivity (Peters 1977).
Eq. (19) has no adjustment for the number of people living in the lake’s
drainage basin, yet we know from physiological studies that animals the size
of human beings release about 0.5 kg P yr
–1
. In other words, each additional
human occupant of the drainage basin has the same effect on phosphorus load
to the lake as an additional 10 ha of undisturbed watershed. Thus, to adjust for
the phosphorus in human sewage in occupied watersheds, one must add
0.5 kg P per occupant to the sum in Eq. (19). If the population uses high phos-
phate detergents, the per capita phosphorus load should be increased to 1 kg
per person per year. These same values allow us to calculate the effect of pro-
posed developments and cottaging on the phosphorus load to a lake. The
effectiveness of the approach has been confirmed in subsequent studies relat-
ing the number of people living in a catchment and phosphorus output
(Mosello et al. 1978, Dillon et al. 1994).
Precept and observation in models of retention. Although we might be
able to predict J and Q, the annual loads of phosphorus and water respec-
tively, we could not use Eq. (17) to predict total phosphorus concentration
without an estimate of the retention coefficient, R.
139A RESEARCH PROGRAM IN HOLISTIC EMPIRICAL ECOLOGY
Geology Dominant land use
Forests Forests + pasture
Igneous 4.7 (0.7–8.8) 10.2 (5.9–16)
Sedimentary 12 (6.7–18) 23.3 (11–37)
Table 12. Phosphorus export coefficients for contrasting bedrock geologies and land
use patterns. Listed values are the means and ranges, in mg m
–2
yr
–1
, of observed
annual losses of phosphorus in all forms expressed per unit area of drainage basin.
(From Dillon and Kirchner 1975)

To predict retention, Kirchner and Dillon (1975) first fit a regression
model to estimates of R from the whole lake phosphorus budgets in Dillon’s
doctoral work and the literature. The model described R as a complex func-
tion of the annual hydrological load expressed per unit of lake area (q
s
= Q/A,
in m yr
–1
). Although the regression fit the data as well as possible, extrapola-
tion beyond the data set suggested that retention would be less than 100% in
lakes with no outflow (q
s
= 0). Since this seemed impossible, they chose a
second, semi-empirical model that was consistent with the expected retention
of 100% when q
s
was zero, at the cost of a slightly poorer fit to the data:
R = 0.426 exp(–0.271q
s
) + 0.574 exp(–0.00949q
s
) (20)
Dillon and Kirchner’s model made sense, but it did so by sacrificing
descriptive power for preconception. Such trade-offs may sometimes be nec-
essary. However, in this case the semi-empirical model (Eq. 20) proved less
effective in predicting retention in lakes with low values of q
s
(Fig. 30).
Ostrofsky (1978) developed a series of empirical models that performed bet-
ter, although they did not predict 100% retention where outflow was zero:
R = 0.201 exp(–0.0425q
s
) + 0.574 exp(–0.00949q
s
) (21)
Ostrofsky succeeded by sacrificing preconception for descriptive power.
Other retention models. Subsequent authors have developed a series of
regressions and other models to handle phosphorus retention, either explicitly
like Eqs. (20) and (21), or implicitly in variations on Eq. (17). For example,
the widely recognized international program of eutrophication research sum-
marized by Richard Vollenweider and Joseph Kerekes (OECD 1982) recom-
mends:
[TP] = (J/Q)/(1 – τ
0.5
) (22)
where τ is the lake turnover time (yr) calculated as lake volume divided by
annual hydrological load (τ = V/Q). In other words, 1 – R = 1/(1 – τ
0.5
). Nürn-
berg (1984) compared a series of possible models and settled on:
R = 15/(18 + q
s
) (23)
Nürnberg also showed that existing models performed poorly in lakes that
developed anoxic deep waters, apparently because anoxia increases the rate
of phosphorus release from the sediments. For such lakes, Eq. (23) should be
corrected by an additional term for internal phosphorus load (J
int
) calculated
as the product of the area of anoxia, the duration of anoxia, and the rate of P
release from anoxic sediments:
[TP] = J(1 – R) + J
int
/Q (24)
140 EMPIRICAL LIMNOLOGY

Holistic tests of the predictions. All the theories predicting the concentra-
tion of phosphorus in lakes would count for nothing if lakes did not respond
to higher phosphorus concentrations by becoming greener, and by developing
other symptoms of eutrophy. When I changed my research directions to pre-
141A RESEARCH PROGRAM IN HOLISTIC EMPIRICAL ECOLOGY
Fig. 30. The failure of precept as a guide in empirical modelling. (A) The model for re-
tention of Kirchner and Dillon (1975) made sense in that it was required to predict
100% retention of phosphorus in lakes with no outflow, but it overestimates retention
(B) where hydrological load is small (Ostrofsky 1978)

dictive limnology, a considerable body of evidence already suggested that
lakes eutrophied in response to phosphorus, but much of that work was open
to criticism and re-interpretations. For example, the classic work of Edmond-
son (1970, 1972, Edmondson and Lehman 1981) detailed changes in Lake
Washington, including phosphorus and chlorophyll reductions, after sewage
was diverted away from the lake. These observations could be interpreted as
the parallel effects of some common, but unmeasured, cause. The bulk of lab-
oratory work suggested that phosphorus was limiting, but there were excep-
tions and the problems of scale hampered the transfer from lab to lake.
Some whole-lake studies appeared to support the role of phosphorus in
the eutrophication of lakes. Schindler’s dramatic experiments with nutrient
addition to lakes showed that massive additions of phosphorus made a lake
green (Schindler 1971, 1974, 1978). In Sweden, early implementation of
phosphorus abatement programs began to yield positive results (Forsberg
1987), and projects elsewhere also showed promise (Sas 1989). However,
fortunes were at risk, both to the public purse in terms of the cost of tertiary
treatment, and to the soap and detergent industry which had invested heavily
in the production of high phosphate detergent, so phosphorus abatement met
active resistance.
I recognized the need for more whole-lake tests of the models, but I was
generally unable to convince my traditional sources of funding that such
money would be well spent. Twenty years ago, Canadian agencies, that is to
say my peers in science, had such faith in the necessity and efficacy of tradi-
tional approaches that they saw little of value in whole-lake experiments and
were appalled at the costs of field surveys that sought patterns among many
lakes. This attitude still prevails at the world’s most important source of
research funds, the National Science Foundation of the United States. As a
result, American ecologists are actively discouraged from empirical, holistic
research in ecology. Since American scientists are among the best supported
and best respected in the field, this antipathy is one of the greatest stumbling-
blocks to the development of effective ecological theory.
Interestingly, applied branches of government in Canada were less reluc-
tant to accept the lessons of our research. Indeed, we were startled at the
speed with which our results were implemented in phosphorus abatement
programs. As dusty academics, we had always believed that government and
politicians ignored our advice out of perversity, ignorance or self-interest.
Instead, the public clamour for action on eutrophication was so loud that
politicians were desperate for applicable solutions. As soon as we had some-
thing concrete to offer, they proved more than ready to listen and to act.
In the past, ecologists had failed to give advice that could be applied. For
example, our traditional advice on eutrophication was essentially to reduce
142 EMPIRICAL LIMNOLOGY

phosphorus loads to zero. Since that goal was impossible, our advice was
ignored. The new models allowed cost-benefit comparisons and showed that
effective abatement was practicable.
I eventually succeeded in finding support for whole-lake experiments,
but only from an unconventional source, The Department of Indian and
Northern Affairs, and then only under the condition that I work in the Cana-
dian Arctic. The department wanted a scientific basis for land use regula-
tions in the north. Since I was now committed to the idea that lakes fit a
common pattern, I saw little immediate need for remote testing, but I was
prepared to make a virtue of necessity by testing the applicability of south-
ern models on lakes lying in permafrost. In this work, we essentially re-
peated Schindler’s whole-lake phosphorus addition experiments, but rather
than raising phosphorus concentration an order of magnitude in a single
lake, we added only enough phosphorus to double the initial phosphorus
concentration of several different lakes. To our delight, the lakes responded
like southern lakes. Our initial surveys showed that the phosphorus-chloro-
phyll relation in these lakes was no different from those established in
southern lakes. Phosphorus addition to some of these waters resulted in
higher phosphorus concentrations, and the chlorophyll levels responded by
increasing as expected (Smith et al. 1984).
The growing school of empirical limnology. According to Kuhn (1962),
programmes in science succeed because they generate more puzzles for sci-
entists to solve. Thus, to predict the greenness of lakes, my group was led to
predict phosphorus concentration of lakes. Those studies in turn led us to
predict retention and the components of the phosphorus and hydrological
budgets, and eventually to test the corpus of existing theory with experiments
on Arctic lakes. It proved a fertile area of research.
The Arctic work was more novel than I had anticipated. The limestone
basins of the area held a surprise: the hydrological budget depended so much
on subsurface flows that Q could not be estimated. As a result, models like
Eq. (17) have limited application in such sites. There is still work to do there,
and at every other stage of the research.
I do not think that our success with the prediction of chlorophyll is a lucky
fluke. I see it as a successful test of the theory that science begins with hol-
istic study and empirical identification of patterns in nature. The same
approach should work elsewhere, on other problems in ecology and limnol-
ogy, and so it has. I will therefore conclude this section with some further
tests of the efficacy of holism and empiricism in producing simple ecological
theories.
Hypolimnetic oxygen deficit. Trout (Salvelinus), whitefish (Coregonus)
and walleye (Stizostedion) are among the most prized game-fish in Canadian
143A RESEARCH PROGRAM IN HOLISTIC EMPIRICAL ECOLOGY

lakes. Because these fishes prefer cold temperatures, they are often confined
to the deeper water of lakes during the summer months. Because they are
adapted to well oxygenated waters, they die if the oxygen concentration falls
below 2 to 3 mg l
–1
, and because eutrophication often results in hypolimnetic
deoxygenation, fish-kills are among the most noticeable and unwelcome
effects of eutrophy. As a result, there was a particular need for models that
predicted oxygen concentrations in lakes. In addition, because anoxia
enhances phosphate release from the sediments, a model to predict the extent
of anoxia should allow improved predictions of internal load. Welch (1974)
and Lasenby (1975) had already developed relations for oxygen concentra-
tions under the ice on frozen lakes, and Jan Barica (1984) had developed
models to predict fish-kills in hypertrophic prairie lakes and sloughs. We still
needed relations for stratified lakes in summer.
At the time, limnologists depended on ideas developed by the founders of
limnology: Thienemann, Strøm and especially Hutchinson. They had devel-
oped three hypotheses about the development of the hypolimnetic oxygen
deficit:
(1) Hypolimnetic oxygen consumption is the result of oxidation of organic
material settling out of the surface waters.
(2) The quantity of this organic material is directly proportional to the rate
of organic production in the surface waters.
(3) The rate of hypolimnetic oxygen consumption is proportional to the
amount of organic matter settling into the hypolimnion.
Since hypolimnetic oxygen consumption depended on the amount of material
settling through the upper surface of the hypolimnion, the rate of hypolim-
netic oxygen consumption was expressed as the average rate of decrease in
the mass of dissolved oxygen under a square meter of the hypolimnetic sur-
face. This is called the areal hypolimnetic oxygen deficit (AHOD, in mg O
2
m
–2
d
–1
).
The three hypotheses listed above were generally accepted in limnology,
and were supported by a series of estimates of hypolimnetic oxygen con-
sumption in lakes of different trophic state by Hutchinson (1938). Unfortu-
nately, no limnologist was able to confirm Hutchinson’s results for other
lakes. Forty years later, Jack Cornett, a graduate student at McGill, succeeded
where other limnologists failed. He showed why no one had been able to
reproduce Hutchinson’s results, he falsified the model under which Hutchin-
son had worked, and he pinpointed the source of this failure as the third of the
listed hypotheses.
Cornett was able to do so for two reasons. He doubted the premises of
Hutchinson’s model, whereas others accepted them without reservation; and
144 EMPIRICAL LIMNOLOGY
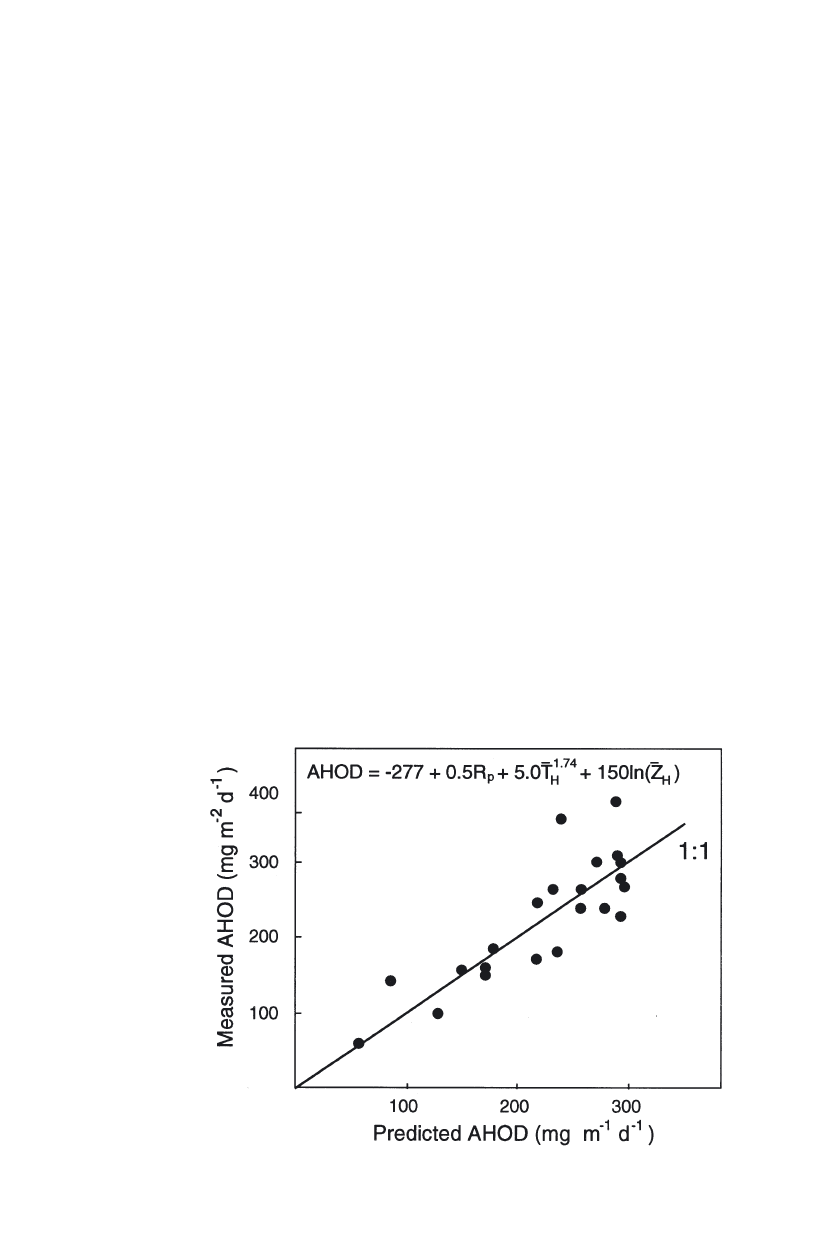
he used the absolute retention of phosphorus (calculated as R × J) as an index
of the amount of organic material falling into the hypolimnion. The second
point is important because it allowed Cornett to estimate organic load in many
more lakes than would otherwise have been possible. Cornett’s model (Fig.
31) shows that the AHOD is a function of the absolute retention of phospho-
rus, the mean temperature of the hypolimnion, and the mean depth of the
hypolimnion.
Cornett did not stop at this point. He recognized that AHOD was not really
what we wanted to know. The hypolimnion is not homogeneous with respect
to oxygen concentration. If we wanted to know what part of the hypolimnion
was anoxic and what part was still habitable by fish, we had to estimate the
volumetric oxygen deficit (VOD) in each stratum. Apparently we had agreed
to look at a substitute variable (AHOD), even if this was not what we wanted
to know, and for many years we were content with this substitution because it
showed what determined hypolimnetic oxygen in principle, even if not in
practice. Eventually, Cornett was able to develop a model which did predict
VOD and therefore could predict the concentrations of oxygen in the water,
the variable in which we are actually interested (Fig. 32, overleaf).
Other phosphorus response models. A number of workers reasoned that if
phosphorus influenced chlorophyll so profoundly, it should affect other com-
145A RESEARCH PROGRAM IN HOLISTIC EMPIRICAL ECOLOGY
Fig. 31. Predicted vs observed estimates of areal hypolimnetic oxygen deficit. R
p
= ab-
solute retention of phosphorus, T
–
H
= mean temperature of the hypolimnion (°C),
Z
–
H
= mean depth of the hypolimnion (m). (From Cornett and Rigler 1979)
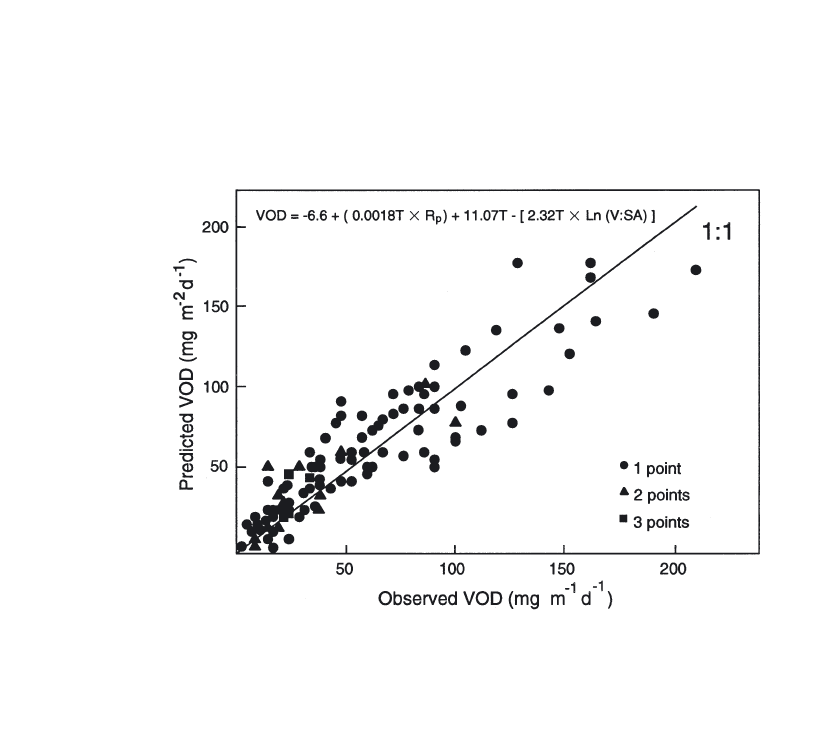
ponents of the lake community too. No doubt some of this work is simply an
analogue of the existing regressions, whereas others are based on a deep con-
viction about how lakes function. The source of the researchers’ beliefs is not
at issue in theory creation. What is noteworthy is that such general success
has been achieved in developing empirical, holistic theories, that the ecologi-
cal characteristics of lakes are more predictable than those of any other com-
munity.
I would need a much larger review than this to do justice to all of pre-
dictive limnology. I can, however, briefly indicate the scope of that work.
Models now exist to predict fish standing stock (Hanson and Leggett 1982),
zooplankton biomass (Hanson and Peters 1984, Pace 1986, McCauley et al.
1988), bacteria (Bird and Kalff 1984), benthos (Rasmussen and Kalff 1987,
Rasmussen 1988), and the distribution and biomass of epiphytes (Cattaneo
1987) and macrophytes (Chambers and Kalff 1985, Duarte and Kalff 1990).
Other models predict primary production (Smith 1979), planktonic respira-
tion (Ahrens and Peters 1991, del Giorgio and Peters 1993) and fish harvest
or yield (Hanson and Leggett 1982, Downing et al. 1988, Godbout and Peters
1988). Antoine Morin developed models to predict the biomass (Morin and
146 EMPIRICAL LIMNOLOGY
Fig. 32. Predicted vs observed volumetric rates of oxygen consumption in different
hypolimnetic strata. T = temperature (°C), R
p
= absolute retention of phosphorus,
V:SA = ratio of stratum volume (m
3
) to sediment area (m
2
) contacting that volume.
(From Cornett 1989)

Peters 1988), production (Morin et al. 1988a) and ingestion (Morin et al.
1988b) of black flies living in streams. Still other general relations exist to
predict the production of aquatic invertebrates and fish (Downing et al. 1988,
Plante and Downing 1989, Morin and Bourassa 1992). Other models predict
characteristics of sediments (Rowan et al. 1992), macrophytes (Chambers and
Kalff 1985, Duarte and Kalff 1990), and periphyton (Cattaneo 1987).
None of these relations are perfect, but none of us expected perfection.
What is far more surprising is the speed and the ease with which empirical
theories were developed for a broad range of limnetic phenomena. This rapid
success gives me hope that many ecological questions can be similarly
resolved, and that useful, credible ecological theory is generally possible. If
so, we may see a generation of applied ecologists who, like physicians and
engineers, have valuable skills to offer, and a generation of ecology texts that
are more similar to engineering handbooks than to philosophical treatises.
Perhaps then humanity will be worthy and capable of the planetary steward-
ship we have assumed.
Summary — A Future for Ecology
In summary, the failure of ecology to produce useful predictions was due
not to the complexity of the subject, but to the complexity of our approach. It
took me twenty years to become dissatisfied with the traditional approach and
to identify an alternative. However, when I changed course, it took less than a
decade to see the results of the new approach put into practice and to see the
rapid success of the approach in addressing a whole range of limnological
phenomena. The process has just begun and I am fully confident that the field
will continue its rapid expansion for many years to come. After all, even the
best of the current models leaves great room for improvement. Eventually,
these patterns may be ready for the next step in science, reductionistic expla-
nation, but for the time-being our needs are such that I am still putting my
faith and my effort in theory creation. I am optimistic that an empirical, holis-
tic approach will be widely embraced by ecologists because society needs
help to preserve itself and its environment, because ecologists want to be use-
ful, and because the empirical, holistic approach is the approach that works.
147SUMMARY —A FUTURE FOR ECOLOGY


X An Education in Science: Evaluation
“The instruction at Edinburgh was altogether by
lectures, and these were intolerably dull...”
Charles Darwin
[The Autobiography of Charles Darwin (1876)]
The thrust of my argument to this point has been that scientific knowledge is
knowledge of a special sort. It uses theory to identify which of the possible
observations are unlikely. The ecology we teach and learn often fails to meet
that criterion, yet there is an alternative. Predictive ecology uses an holistic,
empirical approach to identify pattern in nature. This search for pattern has
been an early step in the development of all scientific theory. Better still, the
identification of pattern is a simple process that uses time-honoured scientific
tools and that lies within the grasp of most working scientists. Even without
genius, we can create a more incisive ecology by thinking about the science
we do.
One element in this introspection is thinking about the way we teach ecol-
ogy and biology. Because one generation of scientists teaches the next, it is
imperative that we consider our present paradigms and methods of teaching,
their past success and their future place in education. We have to teach better
than we were taught or have taught.
On Advising Teachers
Perhaps the most arrogant act a teacher can commit is to advise others how
to teach. What is merely a weakness on the part of a student, parent or admin-
istrator is a sin for the teacher because the teacher alone knows how much
harder it is to teach well than to give good advice. We teachers know how eas-
ily a lesson, conceived like a beautiful dream in the solitude of our office,
becomes a nightmare when presented in class. And although we can dash off
an eloquent description of our philosophy, methods and objectives in an
evening, we rarely, perhaps never, produce a course that actually achieves
those objectives. As a teacher, I should know better than to give advice about
teaching. Nevertheless, I yield to the temptation to do so because I believe

that it is time for a change in the teaching of ecology, of biology, and of sci-
ence. The curriculum we inherited from 19th century Britain is no longer
appropriate to producing the scientists we need.
Even as I set aside my aversion to advice about teaching, I still face daunt-
ing questions like “Is it possible for us to become better science teachers than
we are now?”, “Have we reached the limits imposed by our own ability and
education?”, and “What remains to be said about a subject that has been
worked and reworked by generations of professional educators?” I cannot
answer these questions with authority. When I teach, I merely profess my
beliefs about science to a small group of select students. I therefore have
neither the training nor the experience one expects from a teacher of peda-
gogy. I only have faith — faith that there is always more to be said and faith
that we can always do better. Secondary schools could do a better job of
preparing students for science programs at university, and universities could
do a better job with the students they get. Those of us who are teachers must
ask how this can be achieved.
The Goals of a University Education in Science
To begin, we should ask what we are supposed to be doing for our stu-
dents. In other words, what is the purpose of a university training in science?
I see only two purposes for scientific education, one a question of practical
detail and the other a question of general grasp, but I recognize that these are
stressed differently by different people.
The first purpose is practical. Students expect the university to prepare
them for a career and therefore they want what they call a marketable degree.
Legislators and taxpayers want the university to produce the technicians and
managers necessary to keep a highly industrial, technological society running
smoothly. The objectives of both interest groups can be met by providing stu-
dents with a certain number of specific skills, by giving them the ability to
learn new skills and by instilling in them the confidence to use their abilities.
University teachers sometimes reject the practical demands of students or
society because we think them inconsistent with the goals of a true university.
We therefore spend little time explicitly teaching students the details of doing
things. Instead, we seek to develop a general understanding of the field, pre-
suming that the practical skills needed to solve specific problems will some-
how follow automatically. Unfortunately, the evidence for this coupling of
general grasp with specific capacity is not strong, and many graduates are
frustrated that their general education gives them so little ability to do any-
thing concrete.
150 AN EDUCATION IN SCIENCE: EVALUATION

Our disinterest in teaching people how to do things reflects a long-
standing tradition that science is an aesthetic and contemplative activity, not
a practical one. Many thinkers in ancient Greece eschewed experiment, per-
haps because their manufacturing skills were too rudimentary to test their
sophisticated ideas or because manual labour was appropriate only for slaves
(Macaulay 1852, Russell 1931, Medawar 1984). In the great British universi-
ties, the traditions of scientific thought and discussion grew out of the human-
ities and arts. Applied science and engineering were left to more practical
men outside of academia (like James Watt or Josiah Wedgwood), so that man-
ual skills were undervalued. Until this century, many English scientists were
independently wealthy gentlemen who saw the study of nature as purely intel-
lectual. They devised and used experiments, but the labour and critical tech-
nical skills were provided by unsung technicians (Price 1986). The contem-
plative tradition in science has a long and noble lineage, but it was an error.
We are wrong to treat science only as a matter for the mind.
Universities have always had some pedestrian, practical goals and always
will. Now, when the doing and teaching of science are more expensive than
ever before, universities depend totally on society. University teachers must
therefore serve the state and the students. We must teach them how to do
things. As a bonus, we will find that we also produce better scientists,
because we will produce people who can do their science, not just think and
talk about it.
I do not mean to suggest that the university is nothing but a service insti-
tution. It is and must be a subversive element in society. Its professors should
preach an anti-materialist, anti-establishment, and sometimes anti-religious
doctrine. As the university trains students to be useful tools of society, it
should also tempt them with a vision of total self-indulgence: a life in science,
a life dedicated to inquiry into the workings of the material world. As we train
our students, we should try to change their values, to make them more inter-
ested in the generalities of nature than in the details of making money.
Since our subversion rarely succeeds, university scientists were much
happier teaching the violent rebels of the 1960’s than the docile herds of the
late 70’s and 80’s. Students in the 1960’s seemed so much less interested in
the demands of society, that we teachers thought we had at last succeeded in
our subversive agenda.
Strategies for Teaching
The teacher tries to provide an overview of major findings of the contem-
porary discipline, to initiate the student into the paradigms of science, and to
151STRATEGIES FOR TEACHING

show the student how the findings of the discipline apply to relevant obser-
vations. Some of these strategies will teach students about the theories of the
science, but others can conflict both with that intention and with the expecta-
tions of student and society. Yet, because university teachers rarely specify
either the goals or the strategies of education, they lack explicit guides in the
preparation and presentation of their material. Instead, teachers try to do
everything simultaneously and science courses become self-contradictory,
confused, and confusing. It is therefore a useful exercise to consider what
strategies might achieve the various goals of education.
Empowerment by theory. Theories allow anyone to derive identical de-
ductions from the same, specified information. They have to be used with
care, because they provide the power of science to control our environment
and our lives. However, they are not statements that should be kept out of
the hands of the unwary or the uninitiated (i.e. the students). Instead, an ini-
tiation into the use of theory should be the core of an effective education in
science.
In practice, the application of science is necessarily a question of detail.
It requires a knowledge of theories of very low generality to select appropri-
ately among competing constructs, to choose the appropriate techniques to
monitor a particular situation, to use these techniques well, to assess the
results of this monitoring, and to find alternatives that offset any adverse
effects suggested by assessment. Part of our education should teach this
process through paradigmatic examples of good practice and ecological
success.
Students also need to learn theories of greater generality. Grand theories
provide models for less general theories and contexts for specific actions.
Students must therefore learn the grand theories of science, like the laws of
thermodynamics or gravity. There are also theories of intermediate generality
that link the grand theories to observation and that provide broader, less pre-
cise predictions about the phenomena of nature for non-specialists. The stu-
dent should master some of these intermediate theories too.
A scientific education also transmits the traditions of searching for and
testing theories. It describes some of the successes of theoretical discovery
and application, and outlines the failures of current theory. In Kuhn’s (1962)
phrase, it provides paradigms for future practitioners.
An education in science should teach students how to use some subset of
existing theory, provide them with the skills needed to find other extant theo-
ries, and cultivate their abilities to build, apply, disseminate and judge future
theories. If we taught our students about theory and theories, our graduates
would find they have skills to sell, society would find they have the requisite
technicians, and science would find it has a new generation of able practi-
152 AN EDUCATION IN SCIENCE: EVALUATION

tioners. A student who has had such an education would know some of what
has been done and what remains to do, would know a series of examples of
good scientific practice, would see how techniques condition and interact
with theory, and would have a basic set of theories that allow application of
knowledge to some of the problems of society. Such a student would have
been empowered by his or her scientific education.
Understanding through explanation. Scientific knowledge is predictive
and theories are the constructs that make predictions. I would like to be able
to claim that an education in ecology is based on theory, but no such claim is
justified.
If one searches the indices and tables of contents of current ecology texts
for references to “theory”, one is struck with how rarely the word is used.
When it is used, the word almost invariably denotes a highly academic con-
struct which makes few, if any, predictions. Most ecology courses, texts and
teachers try to describe the contemporary science, and therefore do not stress
the role of prediction and theory. Because ecologists often do not recognize
the central role of theory in science, we do not yet have the theories we need,
we do not teach the few theories we have, and our students would be unable
or unwilling to learn them if we did. In short, courses teach what ecology is,
not what it does or should be.
Many ecology teachers have adopted a strategy designed to produce a
sense of understanding, rather than a mastery of ecological theory. In provid-
ing a sense of understanding, science makes us feel “at home” in the universe
and so performs a role similar to that of religion, art, and an assortment of
other human activities. These activities succeed by giving us the feeling that
we understand and control events in our lives, even if this is not so. In reli-
gion, the rituals associated with the winter solstice appear to bring longer
days and, eventually, spring. In art, the irrationality of poetry may seem to
explain events and feelings we do not understand at all, like love, beauty,
laughter, and grief. And in science, an educated student can expound at length
about the causes of the weather, the impact of pollution, and the future of
AIDS, even though the phenomena may be totally unpredictable. In all fields,
the importance of understanding is not whether we actually control events,
because we certainly do not, but to make us feel less at the mercy of unknown
powers and forces.
To promote understanding, teachers try to explain relevant aspects of the
universe using the constructs of their discipline. In principle, these explana-
tions should demonstrate that a given observation could have been pre-
dicted, and thus that the observation was an instance that could have been
deduced from a more general theory (Hempel 1962). In practice, the
elements of explanation are a hodge-podge (Peters 1991a). By offering
153STRATEGIES FOR TEACHING

alternatives to predictive theory, the search for understanding usurps the
rightful place of whatever theories we have, and hides our ignorance with
word-play.
Putative explanations based on non-predictive statements are no more than
post hoc rationalizations. Because the human mind can explain any finite set
of observations in many ways, such explanations are always possible and
plausible. Given the advantage of hindsight, post hoc explanations can fit the
available facts exactly, whereas predictions from theory are unspecific state-
ments of probability. So long as we only compare explanations for data that
have already been observed, post hoc rationalizations appear more precise
and attractive than scientific explanations.
To distinguish post hoc rationalization from scientific explanation, one
must ask if any observation would have invalidated the explanation. Because
post hoc rationalizations are specific and complex, they need only explain
what occurred. Alternative observations are easily explained away by differ-
ences in the details of each case. In contrast, scientific theories not only pre-
dict some observations, they also prohibit others. The theory would be
falsified if these other observations were made. This asymmetry has the
regrettable consequence that post hoc rationalizations are difficult to remove
from the field, so they tend to accumulate in the literature. Theories, and
explanations based on theories, can be falsified and forgotten. In ecology,
non-scientific explanations provide students with poor examples, and hide
the fact that we can predict relatively little and then only imperfectly. This is
most regrettable because, if we recognized the extent of our scientific igno-
rance, we would also see the many opportunities for improvement that ecol-
ogy offers.
We likely find non-scientific understanding seductive, because we are
unwilling to admit how little we control our environment. If we accepted pre-
dictability as the criterion for scientific knowledge, we would realize that
most of the decisions that govern our lives are not scientifically justified.
Economics is a predictive swamp, politics a scientific quagmire, social inter-
action a bog of intuition, and so forth. If we recognized the limits of our pre-
dictive and manipulative power, our already deep sense of helplessness might
become unbearable.
The explanatory role of understanding may reflect a deeply seated human
need, but that feeling must not hide the importance of predictive power. An
obsession with understanding draws potential researchers away from science,
and the desire to explain makes scientists reluctant to accept Popper’s pre-
dictability criterion as the demarcation between science and non-science.
Instead of confronting scientific short-comings, educators frequently disguise
them with words of explanation and a sense of understanding. This preserves
154 AN EDUCATION IN SCIENCE: EVALUATION

the status quo and social serenity, but confounds the advance of science
because it obscures the problems that scientists should address.
Paradigmatic indoctrination. A common strategy in teaching is to
indoctrinate the students to think and act like their teachers. This is the point
where we teach as to “intending professionals” (Barzun 1964) and where stu-
dents are introduced to the over-arching paradigms of normal science (Kuhn
1962). Teachers know that future researchers will not live or work in a vac-
uum. Would-be scientists must be recognized by the leaders in the field and
by peers, or they will never have a chance to practice science. As in art or in
any creative field, a would-be innovator must play by the rules that govern the
community. Scientific innovators must address questions that are of interest
to their community, they must design their experiments in the appropriate
manner, and they must analyze the data by accepted methods, even if this
means going to extremes, like applying advanced statistical tests where none
is necessary. Fledgling scientists must publish in accepted journals, even
though these are read by only a handful of professionals, and they must write
in the accepted style, even though that style is pompous, dull and nearly
incomprehensible. New scientists must be indoctrinated to do all these things
or their work will not receive a fair hearing.
If we want our students to succeed in science, part of our effort must be to
teach them to conform. However, conformity holds the danger of stagnation.
When a discipline can no longer meet new demands placed on it by society
and by its members, the field is in crisis. New solutions and approaches out-
side the normal paradigms are sought, and eventually a new paradigm may be
found to treat contemporary concerns. Progressive science therefore needs
the occasional revolution (Kuhn 1962).
I believe that environmental degradation has revealed the inadequacy of
traditional ecological theory. Ecology needs a revolution, but the processes of
critical assessment and radicalization that precede revolution are frustrated by
indoctrination into contemporary ecological paradigms. At the end of their
education, too many students simply accept the foundations of their science,
and the only problems they can find to study are minor elaborations of current
paradigms. We teach people to be normal scientists, but we may need a revo-
lution to meet the challenges of contemporary ecology.
Disciplinary description. A description of findings in the discipline is a
part of all strategies: teaching theory, developing understanding, inculcating
paradigms, or promoting revolution. However, theory is only part of descrip-
tion, and in ecology usually a minor part. Even upper level courses may
describe contemporary science as a body of incontrovertible concepts, logical
truths, and law-like facts. Students are encouraged to believe, rather than to
question the corpus of science. As a result, even senior undergraduates and
155STRATEGIES FOR TEACHING

graduate students see science as virtually complete and consequently rather
uninteresting. To the converted, contemporary science seems unlikely to
change.
An Evaluation of Teaching in Biology and Ecology
To summarize the chapter to this point, I have suggested that teachers
should strive to prepare students for careers, to prepare technicians for the
state, and to infect some young minds with the passion for science. If we are
doing our jobs well, students graduating with a B.Sc. should be fluent in the
use of a body of scientific theory and method, and they should be confident in
their ability to learn more skills and theories. Those who go on to research
need these same attributes, but they also need an appetite for scientific
thought and investigation.
A simple example will show how well I think we are achieving the practi-
cal goals of higher education in science. I recently had to hire a technician
and my advertisement attracted five graduates from the undergraduate pro-
gram in biology at my university. To select among them, I thought to use a test
to determine if the applicant could follow routine analytical procedures. I
asked each applicant how he or she would prepare a one molar solution of
sodium chloride and how they would dilute that stock to 0.1 M. I thought this
task would present no difficulty because our undergraduates are required to
complete several courses in chemistry, and the applicants were allowed to use
any books they wished. What astounded me was not just that none could
answer the questions directly, but that none could describe how they might go
about finding out how to answer them. Apparently, they could not apply what
they had learned and were so unconfident in their abilities that they could not
teach themselves either old or new material. How will they meet the chal-
lenges of the current marketplace, much less of the next millennium where
they would spend most of their lives?
Nurture or nature. If our students are not as capable as we would like, we
can blame their genetics (them) or their environment (us). The only fruitful
approach is to blame ourselves.
The material society sends to be educated must be basically sound. During
rare historical periods of exceptional intellectual achievement, small popula-
tions have produced many more masters and geniuses than numbers alone
would suggest possible. One need only compare the architecture and sculp-
ture of Periclean Athens, the painting of renaissance Italy, the literature of
Elizabethan England, or the music of 19th century Germany with the prod-
ucts of our age. These societies contained only a few tens of thousands of
156 AN EDUCATION IN SCIENCE: EVALUATION

potential contributors, many of whom lived in poverty, ignorance and squalor
compared to us. Yet they achieved much more than the well-fed, leisured
millions who make up contemporary mass society. If we cannot succeed in
educating scientists, it is more likely because we, as a society, spoil the mate-
rial we have, than because we receive spoiled material.
Even a few hours with a pre-school or kindergarten class is enough to
show that most young children have ample wit and intelligence. Even later,
after the schools have done their worst, they still deliver superior students to
the universities. For example, Harmon (1961) analyzed the secondary school
IQ’s of eventual winners of doctoral degrees in different fields (Table 13), as
revealed by U.S. Army General Classification Test (AGCT) scores. These
results show that we can be elitist in thinking about education.
Both lines of evidence, the usually untapped potential of humanity and the
demonstrated intelligence of our students, imply that the failure of a science
education is more likely to be institutional. High IQ’s and native ability are
not enough. Indeed, most of our doctorates in science do not succeed in their
discipline. De Solla Price (1986) has shown that of all the contributors to a
field in a given year, a fifth never publish again in the same area and a further
third never publish again in any field. More than half the contributors to a sci-
ence do so only once and then drop out. Only a fifth of the initial contributors
become the core of researchers who are major contributors to the science over
the longer term. We are not using our material well.
The problem with textbooks. If biologists are uncertain about the nature
of science, they cannot be expected to direct students effectively. This uncer-
tainty is echoed in the cursory descriptions of the nature of science offered in
introductory textbooks (Chapter I), but is also apparent in the body of the
textbooks, in courses and in more advanced monographs. When teachers are
157AN EVALUATION OF TEACHING IN BIOLOGY AND ECOLOGY
Discipline IQ
Physics 140
Mathematics 138
Engineering 135
Geology 133
Arts & Humanities 132
Chemistry 132
Biology 126
Education 123
Table 13. IQ scores as recorded in AGCT tests of high-school students who eventually
earned doctoral degrees in different fields. (From Harmon 1961)

uncertain about the field, they are unlikely to deliver clear messages about
what is important to their students.
Evidence of the teachers’ uncertainty about what is important can be seen
in the vast welter of information that typifies contemporary texts. Most intro-
ductory textbooks in biology or ecology are huge undertakings stretching
over hundreds of oversized pages. Even these vast texts soon prove insuffi-
cient to contain all our important findings so new, even larger editions appear
at regular intervals (Table 14).
No instructor can seriously suppose that students will absorb more than a
tiny fraction of this material. The texts therefore cannot teach, and they are
scarcely authoritative enough to be reference works. They serve mainly as
samplers of biological thought, providing a place for most major currents in
the discipline. Publishers approve of this format because such a text will
likely touch on the special interests of the professors who select the texts. The
158 AN EDUCATION IN SCIENCE: EVALUATION
Biology texts
H. Curtis y 1968 1979 1983 1989
n 854 1043 1159 1192
W. T. Keeton y 1967 1972 1980 1986
n 955 888 1080 1175
C. A. Villee y 1960 1962 1972 1977 1985 1989
n 615 625 915 980 1206 1412
P. B. Weisz y 1959 1963 1967 1971 1982
n 796 786 886 656 1009
Ecology texts
C. E. Krebs y 1972 1978 1985
n 694 678 800
E. P. Odum y 1953 1959 1971
n 384 546 574
E. R. Pianka y 1973 1978 1983 1988
n 356 397 416 468
R. Ricklefs y 1973 1979 1990
n 861 966 896
Table 14. The growth of textbooks in ecology and biology (as measured by number of
pages, n) with successive editions (as indicated by the year of copyright, y). Page sizes
also tend to increase in subsequent editions, so this table under-represents growth in the
contents of these texts. Names are those of the authors. Editions were selected accord-
ing to availability in the McGill University library system

teacher, recognizing the impossibility of teaching all of contemporary biol-
ogy (or ecology or limnology), can assuage the inevitable sense of inade-
quacy with the thought that the subject is dealt with somewhere in the text.
Interested students could always pursue the topic there, but the indigestibility
of the material must discourage all but the most dedicated (or insensitive).
Texts do not have to be so comprehensive. The best overview I have ever
read of then contemporary biology was The Ideas of Biology, a pocket-sized
paperback less than 200 pages long (Bonner 1962). T. H. Huxley (1880)
developed a text for biology that dealt only with a single model animal, The
Crayfish. Both authors succeeded because they ignored the imperative to pre-
sent everything, and chose to display their ideas clearly and effectively with
highly selected examples. The student could then use these ideas to organize
other facts and other theories. Contemporary textbooks may begin with simi-
lar aims, but they become cluttered by a seemingly inevitable accretion of
biological detail. Each new edition becomes more bloated as the authors add
information to bolster and illustrate their original conception of the science
(Table 14). Unfortunately the result obscures that conception. The secret of
great teaching lies as much in what is left out, as what is left in.
The problem with courses. The swelling texts are only indicators of
teachers’ confusion about biology. The same process occurs in the design of
university courses, as I can illustrate with another personal example.
Some years ago, the government of Ontario decided that the present biol-
ogy course for senior secondary school students was old-fashioned and that it
had to be jazzed up to appeal to modern students. A committee was formed
and a number of zoologists were invited to provide ideas. We talked and
argued all evening. Finally, a good friend of mine announced that he had the
solution and began to outline a new course. Other committee members added
material that was dear to their hearts, and eventually a new biology course
was developed. This is a quite typical ontogeny for an introductory course,
and I suspect that it has been repeated many times in both secondary schools
and universities. Nevertheless, the final product was a recipe for disaster.
The “new course” my friend helped design for high school students was
actually the course he was then giving to first-year university students. In
proposing that a university course be offered to high school students, he over-
looked the fact that university students had only 18 classroom hours a week,
so they had much more time to think about the subject than students spending
almost 30 hours a week in secondary school. Moreover, he ignored the reality
that a university professor might have only 5 contact hours a week, whereas
colleagues in secondary school had 25. In addition, my friend was a highly
trained and brilliant biologist who later became president of a major univer-
sity. This combination of native ability, professional training, and reduced
159AN EVALUATION OF TEACHING IN BIOLOGY AND ECOLOGY

teaching load should have afforded him greater success in his course than
might reasonably be expected of the average high-school teacher.
The real situation was even worse. Because the two of us were working
very closely, I attended many of his lectures, and I repeatedly found he had
serious difficulty understanding some of the concepts he was trying to teach.
Therefore the new course began with subject matter that a good professor
found difficult, yet the course would almost certainly be taught by less com-
petent teachers to more harried students. I think it extremely unlikely that
most students would find that such a course would pique their interest in
science.
My moral is that we cannot teach everything. There is too much for one
course or one career. Instead, we must use a limited set of materials and
examples to show our students how to learn and how to teach themselves.
Repercussions for graduate training. Not surprisingly, the lack of focus
that characterizes undergraduate texts and courses reappears in the research
proposals that students make soon after entering graduate school, and in the
theses they write at the end of their formal education. Students who are not
trained to cherish scientific theory are less capable of formulating testable
hypotheses for their own research. Too often, students set off “to assess the
impact” of some perturbation, “to determine the importance” of some pro-
cess, “to understand the role” of some organism or “to shed light” on a
phenomenon. These woolly generalities provide little direction and convey
little meaning. They hide an unwillingness to specify what variables will be
measured and what responses are expected. They show that one has not
thought about what one can and should do as a scientist.
The challenge for graduate students is therefore to rise above the general-
ities of undergraduate education by identifying a much more specific, imme-
diate goal for graduate research inside a larger context, as an early step to a
long-term goal. This process requires that the generalities be replaced with
highly simplified hypotheses about the relations between operationally
defined variables. It is however not enough that a prediction be made about
some phenomenon; the relations and predictions should also be scientifically
or societally relevant.
So little is known about so much that there should never be a dearth of
good scientific projects. If there seems to be, it is because students are poorly
prepared for the process of hypothesis formulation and testing. The store of
proper paradigmatic examples is low, the testable implications of the science
they know are few, and their graduate instruction, the antidote to an unsatis-
factory undergraduate experience, is weak.
Graduate students in ecology too rarely recognize the futility of a science
that cannot predict. Their long experience with non-scientific “explanation”
160 AN EDUCATION IN SCIENCE: EVALUATION

has crushed the curiosity, doubt and frustration with ignorance that should
give impetus to science. They see their task as the development of discursive
explanations for an accepted set of observations. Instead they should identify
an hypothesis that could have predicted those observations and set out to test
its predictions. Others assume causal relations in nature and set out to prove
the validity of their assumptions, rather than testing what the relation pre-
dicts. Still others study things simply because they were previously unde-
scribed. Small wonder if after five years of studying inward-looking, often
dead-end projects, the graduated Ph.D. leaves the field.
A lesson from the literature. The problem of trying to do too much is not
limited to formal education. Some years ago, I had occasion to read Alfred
North Whitehead. I found it a humbling experience. Like many great
thinkers, Whitehead had recorded a very large number of interesting ideas.
There was no need for the ideas to be correct, and because correct ideas are
very rare, most of them were not. However, great intellectuals, like White-
head, are arrogant people (otherwise they would not have the confidence to
spend so much time writing), so they produce a vast volume of writings. To
do justice to this material would take years of study, and so almost all readers
are inadequate to the task put to them by the world’s leading intellectuals.
The fault does not lie with the busy reader alone. Great writers have writ-
ten far too much. The same idea may be flogged in different guises in dozens
of books. Padding appears everywhere. The great thinker may develop a liter-
ary style that is at times outstandingly good, but at other times obscure. And
the reader, overawed by the writer and by the obscurity, style and quantity of
the material, is simply incapable of sorting out the useful bits. Consequently,
reading becomes superficial, works are labelled thought-provoking (although
there is rarely time to pursue the provoked thoughts), the writer’s fame
spreads, and more, ordinary mortals feel compelled to read the great man’s
works. Even the writer may be fooled and feel encouraged to broaden his
field. For example, although I was struck by Whitehead’s earlier work, I find
his later, more popular, philosophical writings to be so nearly meaningless as
to be incomprehensible. Eventually, even Whitehead exceeded his ability.
This raises a problem. How much time should the would-be reader devote
to the work of a man like Whitehead? A year would surely be far too little, yet
there are many others who merit study. What is the busy scientist or student
to do? The only answer must be to use teachers who can interpret the work for
other professionals, as Magee (1973) and Pera (1980) interpreted Popper.
Science requires incisive teachers at all levels.
161AN EVALUATION OF TEACHING IN BIOLOGY AND ECOLOGY


XI An Education in Science: Prescriptions
“It is this union of passionate interest in the detailed
facts, with equal devotion to abstract generalization
which forms the novelty in our present society....
This balance of mind has now become part of the
tradition which infects cultivated thought. It is the
salt which keeps life sweet. The main business of the
universities is to transmit this tradition.”
A. N. Whitehead
[Science and the Modern
World (1925)]
In the previous chapter, I described the roles we expect our education system
to play, I gave a simple observation suggesting that there is scope for im-
provement, I identified shortcomings at every level, and I suggested that, in
teaching science, the critical issue is our failure to appreciate the nature of
theory.
The interesting question is “How can we do better?” I am going to
approach this question in a way that is somewhat foreign to professional edu-
cators, but more appropriate to a working scientist and teacher. I will ask
“What makes a good scientist?” and “Can we modify our approach to educa-
tion in any way so as to produce more or better science?” This has the distinct
advantage of bringing the discussion closer to my own experience as an under-
graduate instructor, a graduate supervisor, and a university administrator.
The Undergraduate Program
Let me premise this discussion with the observation that the public’s love
affair with universities has grown cold. University professors will have to
grow accustomed to more meagre moral support, less lavish budgets, and
poorer facilities. Our traditional teaching methods will have to change.
We should not expect that any substantial fraction of the teaching staff at
a university would agree on a particular solution or collective action to these
challenges. Professors are free-thinking, angular individuals who are too con-
vinced of their own righteousness to be much influenced by someone else’s

prescriptions for better teaching. This is how it should be. It would be equally
vain to hope for an institutional response to our difficulties. The unhappy
mass of university educators are pulling the administration in a hundred dif-
ferent directions. Experience at two universities has taught me the virtual
impossibility of an imposed, general solution to the problems of university
education.
The only easy modifications that university teachers can impose on educa-
tion involve their own courses. Chapter X indicated where some changes are
needed, and this chapter indicates some that I try to apply in my own teach-
ing. There is no point in advocating the general introduction of these sugges-
tions, so I offer them for other teachers to use or ignore as they see fit.
The problem of confidence. Over a century ago, Francis Galton (1875)
showed that almost half the members of a group of eminent scientists were
either the eldest or only child in their family (Table 15). This remarkable
observation has since been confirmed and extended. A very high percentage
of our best scientists were the eldest or only child (Table 16), they lost one
parent by death (Table 17) or divorce, and they were lonely, rather asocial
children (Roe 1953). These observations should not suggest that, to produce
researchers, we should identify bright children before they reach 10 years of
age and isolate them from their parents. No one advocates such extreme mea-
sures because they would be both morally wrong and ineffective. Roe (1953)
makes the point that about 25% of homeless men also lost a parent before the
age of 10. Most bright orphans do not achieve scientific fame.
The critical characteristic of the bright orphans who do develop into
researchers is that they are very self-confident and self-reliant with respect to
their interactions with the material world. (Their social interactions are much
less successful, but perhaps that is more incidental to their scientific achieve-
164 AN EDUCATION IN SCIENCE: PRESCRIPTIONS
Birth order Number of scientists
Only child 22
First born 26
Middle children 36
Last born 15
Total 99
Table 15. The birth order of eminent English scientists in the 19th century. The families
of these 99 individuals were large, averaging 6.3 children per family, so the chance of
the eminent scientist being the first or only child was much less than the observed
frequency of almost 50%. (From Galton 1875)

ment.) Family circumstances develop in them the belief that they can do and
understand things. This belief is critical to success as a scientist.
For those bright children who are not fortunate enough to be orphaned
before the age of 10, we teachers should help develop this same confidence.
Present science curricula may be intended to achieve that goal, but unfortu-
nately, they are rarely designed in a way that will achieve their intentions. The
main problem is that our science curricula expect far too much of the stu-
dents. They try to be too good. Unrealistically high goals teach students that
they cannot do science, and massive texts show that there is too much mate-
rial to master. Students are therefore trained to pass exams by learning lec-
tures, so they learn to rely on authority rather than reflection, reading and
experience. As a result, university courses rarely teach students to use or to
improve what they have memorized, and certainly do not encourage students
to be confident in their ability to criticize, to use or to do science. The great-
165THE UNDERGRADUATE PROGRAM
Birth order Number of scientists
139
213
33
43
52
62
72
Total 64
Table 16. Birth order of 64 eminent American scientists of the 20th century. Fifteen
only children are included among the first born. (From Roe 1953)
Future field Age at parent’s loss: Total
< 10 yr > 10 yr
Biologist 25 0 25
Physicist 13 9 22
Social scientist 9 18 27
Total 15 9 25
Table 17. Percentage of future eminent scientists who lost a parent in childhood through
death. The expected values for college students would be 6.3% before 10 years of age
and about 10% in total. (From Roe 1953)

est weakness of our present system is that it does not give students confidence
in their abilities.
De-enrichment and dis-integration. Universities and departments like to
present comprehensive programs and integrated courses, perhaps because
such organization implies a mastery of the material. Unfortunately such an
approach misrepresents biology and ecology. Neither discipline consists of
monolithic sets of theories. There is no over-arching theory of great general-
ity that ties together all of modern biology or ecology in the sense that every
theory is a refinement or specification of the general theory. If we insist on
teaching as if there were a single science of biology or ecology, or a central
core of crucial facts, we misinform the students about biology and insure our
own failure as teachers of science. There is instead a heterogeneous collection
of hypotheses with different scopes, variables, formats, domains, and suc-
cesses. Indeed, there are so many of these theories of low generality that they
could never fit into a single degree.
If we wish to give students a realistic view of our science we must teach its
theoretical structure. This requires that we give them a dis-integrated view,
one without a unifying central theory. They must learn that theories in plant
ecology can be falsified or confirmed without impinging on theories of hor-
monal balance, that one can be a good physiologist without mastering popu-
lation genetics, and that success in eutrophication control does not insure
equal success in controlling macrophytes or predicting the yields of terrestrial
herbivores to hunters.
Since we cannot expect students to see the generalities of science if they
are constantly harried and bewildered with the particularities of its sub-
disciplines, students with intelligence and an interest in science require a
factually impoverished or de-enriched program. Teachers might try to cover
one-fifth of the subject matter in most contemporary courses, but ensure that
everything taught in one course reinforces and uses everything taught in other
courses. This would allow students time to contemplate the nature of science
in general and their material in particular. Not all students would do so, but at
least those who wanted to could. The others would be no worse off than they
are already. A de-enriched biology course would cover very little factual
material, but it would show students how to apply knowledge from other
courses and it could build their confidence, graduating students who think “I
can do biology”.
I would teach a few aspects of some homely subject, like limnology or
physiological ecology. These are beautiful subjects with which to introduce
biology because so many aspects can be quantified, and because they depend
on the mathematics, physics and chemistry the students have already learned.
In the laboratories, I would concentrate on a few basic skills in mensuration,
166 AN EDUCATION IN SCIENCE: PRESCRIPTIONS

analysis and handling of biological material using phenomena that are sus-
ceptible to study. Students can easily design and build a water bottle to sam-
ple at depth or a Secchi disc to test general theories. They can build and use
respirometers to measure oxygen consumption by poikilotherms at different
temperatures and levels of activity. They can design and build the equipment
necessary to investigate factors limiting the growth of algae. They can learn
to do biology, and thereby learn to do science. If they do not do so, they will
be lost regardless of the number of facts, concepts and observations they
accumulate. Having done so, they can learn most details about the biology
they need for their careers on their own.
I would also include a hefty dose of the history and philosophy of science
in university training. That material is essential to give students the breadth
and vision to appreciate their science. Philosophy can be a tool to help stu-
dents find optimal strategies in learning and research. It is also a handy
weapon for students to defend themselves against the inevitable attacks from
traditionalists.
There are topics which I would no longer teach. For example, the theory of
evolution by natural selection is a subject that still confuses professionals,
and one that cannot be related to most of the physics, chemistry or mathe-
matics that the students are learning or have learned. It is unlikely that uni-
versity courses will save many fundamentalist students from their ignorance,
and open-minded students can learn most of what they need from a host of
splendid books on evolution, or even from the public press and electronic
media. Above all, neither scientist nor student can do much with the theory
of evolution by natural selection because it makes so few predictions. This
“theory” may satisfy the students’ urge to understand their world in some
emotional, visceral way, but such a sensation is not science. Teaching stu-
dents to seek that form of explanatory gratification is a disservice to their
training as scientists.
I would also avoid some of the exciting new fields, like biochemistry and
biophysics. If they are to be mastered, these fields require a background in
mathematics, physics and chemistry that few biology students have as under-
graduates. In any case, evolution, biochemistry and biophysics will still be
taught by the many professors who will remain with the old curriculum, so
there is no need for me to repeat that material.
A de-enriched and dis-integrated program will impose difficult decisions
on the teacher. The material chosen for the course must be carefully selected,
but no general theory exists to identify relevant material within the sea of bio-
logical detail. The criteria could not be those used by the writers of text-
books, trying to touch all the bases of a huge and complicated field. Nor could
the criteria be those used in so much of contemporary ecology, tying concep-
167THE UNDERGRADUATE PROGRAM

tual argument and extreme detail together with a sense of understanding.
Training different ecologists will require different criteria of biological rele-
vance.
Hierarchical themes for undergraduate education. In the absence of
central theory, relevance in biology or ecology depends on the scientific
demands of society and pedagogy. Material for a course can be selected
because society is interested; for example, we might teach about diseases,
conservation, pollution control, plant production or human population
growth. Material may also be selected because it teaches an effective way to
do science. If we were less concerned to detail all the fruits of scientific inves-
tigation, we might introduce students to particularly creative modes of
thought, to recurrent patterns in the development of theory, to effective criti-
cism of scientific ideas, to effective tests of theories, and to appropriate
responses to both the confirmation and falsification of theory in such tests.
Students would therefore face a series of different examples of scientific and
societal problems that have been addressed, more or less successfully, at dif-
ferent scales and levels of biology.
Such an approach necessitates “hierarchical thinking” (Allen and Starr
1982, O’Neil et al. 1986). The students should learn that similar problems
expressed at different scales may yield to different, even contradictory, solu-
tions. For example, annual mean concentrations of zooplankton and algae are
positively related across communities (McCauley and Kalff 1981, McCauley
et al. 1988), but inversely related in microcosms (Novales et al. 1993) and
over a single season in a single lake (Lampert et al. 1986, Sommer et al.
1986). Phosphorus may be important in lakes (Peters 1986), but marine sys-
tems may depend on nitrogen or iron (Price et al. 1991). An effective course
should therefore draw examples from a range of phenomena and biological
levels to illustrate differences and inconsistencies in our knowledge (Fig. 33).
Within one hierarchy, the student should be introduced to theories of high
generality, which allow predictions of a crude sort but at a very general level,
and theories of low generality that allow more precise predictions about ques-
tions of specific interest. For example, one can make crude estimates of pro-
duction per unit biomass from body size alone for all animals and protozoans,
but if the question is restricted to a single species or population, more precise
and often more complicated relations may be necessary.
The theories of production in Fig. 33 predict similar variables, but at dif-
ferent levels of generality. They coexist because they have different domains
of application. They can be independently falsified or confirmed because they
fall at different points in a hierarchically differentiated science. Similarly,
theories about respiration or ingestion can be resolved at different levels,
again more or less independently of parallel hierarchies. For example,
168 AN EDUCATION IN SCIENCE: PRESCRIPTIONS

169THE UNDERGRADUATE PROGRAM
Fig. 33. One system of theoretical hierarchy that may be used to structure biological
education. The figure shows the relations among theories dealing with productivity, at
various levels of generality, predictive power, and specificity, the relationship of these
theories to other hierarchies dealing with other phenomena, and the relation of all these
hierarchies to still more general theories, like the laws of thermodynamics. I, R, D E,
and P (ingestion, respiration, defecation, excretion and production) are the components
of the balanced growth equation. In the theories of lower generality, B is biomass in g
m
–2
, W is individual body mass in g, T is mean temperature, P is production and C is
fish catch, both in g yr
–1
. Subscripts indicate if the units of mass are wet (w) or dry (d)

production may be predictable from size, but theories of respiration could be
based on temperature and those for ingestion on food concentration, all oper-
ating under the general restrictions set by the laws of thermodynamics, but
otherwise quite free to use different variables, analytical techniques, and
theoretical structures.
If this material is to be taught, the links among the examples must derive
from an explicit consideration of the qualities of good scientific practice. Stu-
dents should learn about the nature of science, the place of creativity and crit-
icism, the role of theory, and the purposes of observations and tests. Particu-
lar examples of success and failure should be discussed and a framework
of interpretation provided.
A theoretical typology. One format with which to organize such a dispar-
ate set of ecological or biological theories invokes a typology of theories. For
illustration, I have identified four basic types of successful theory, although
there may be others and alternate typologies exist (e.g. Orians et al. 1986).
(1) Simple interpolative theories simply assume that a phenomenon mea-
sured at two or more points persists at intervening points. A common type of
interpolative theory in ecology is the species distribution map. If a species is
recorded at two sites we posit that it will be found at intermediate positions,
provided physical-chemical conditions are similar. In fact, we often use the
absence of a species as the first indication of subtle physical or chemical
change.
(2) Limiting factor, mass balance and inter-conversion theories form a
complex group of interrelated, but simple theories based largely on consis-
tencies in the stoichiometry of living systems and the empirical observation
that matter and energy are neither created nor destroyed in biological reac-
tions. These theories are the basis for all the most important predictions that
ecologists have made. For example, such theories allowed Vollenweider,
Dillon, and Cornett to make some very quantitative predictions about the
future of lakes (Chapter IX).
(3) Empirical holistic theories form the biggest and most diverse class of
all. It is less a distinct set than it is one end of a spectrum of all theories. Such
theories include predictions about the performance of an organism based
on its size (Peters 1983), predictions about species number based on evapo-
transpiration (Currie and Paquin 1987, Currie 1991), predictions of litterfall
based on latitude (Bray and Gorham 1964, Lonsdale 1988), and predictions of
bioconcentration factor and effective lethal concentrations based on the rela-
tive solubilities of organic contaminants in water and an organic solvent
(Verschueren 1983, Hermens 1986, McCarty 1987).
Ecologists frequently underestimate the value of empirical theories. Often
we denigrate them as “black box theories” or “engineering”. At worst, we
170 AN EDUCATION IN SCIENCE: PRESCRIPTIONS

ignore them or believe they are not theories at all. This behaviour is self-
defeating because if we were to look over the history of successful branches
of science, we would discover that the most elegant analytical theories began
their lives as humble empiricisms.
(4) Analytical theories lie at the other extreme of this spectrum. The differ-
ence between empirical, holistic theories and analytical theories is that the for-
mer merely predict, whereas the latter also give us the impression we know
why the theories work. An analytical theory predicts and makes us feel at home
by convincing us that we understand the system (Lehman 1986b). Analytical
theories are usually, but not necessarily always, expressed in terms of the sub-
systems of the system. Sometimes it merely invokes fictitious state variables,
such as gravity, magnetism or even animal magnetism. Most scientists aspire
to produce such theories, but there is a time and place for analytical theories.
The history of science shows that they follow empirical, holistic theories.
I see no gain in imposing my views on others, but I can urge that all of us
who teach an approach to science do so explicitly. If our differences are
exposed fairly and conscientiously, students would have a better grasp of both
the strengths and weaknesses of science. Freed of popular misconceptions,
they would be more confident in their ability to perform as well as their teach-
ers, more willing to cast aside theories and constructs that have outlived their
usefulness, and more prepared to accept the responsibility of creating new
knowledge. They might even discover the real joy and freedom of research.
Graduate Education
One should not do science because it offers a socially acceptable career
with adequate pay and interesting fringe benefits, but because something in
one’s nature compels one to do so. Such a student will find graduate school a
reward in itself and the supervisor should ensure that this is possible.
Many scientists remember graduate school as a golden time when they had
more intellectual freedom, more time to reflect, and fewer extraneous cares
than at any other point in their lives. To preserve as much of this experience
as possible, one must settle on a strategy for graduate study that allows stu-
dents the freedom to do science, and is likely to offer them the chance to con-
tinue to do so after graduation.
The importance of role models. One way to develop theories about effec-
tive education is to look for pattern in the training of highly successful scien-
tists. Galton (1875) provided one such pattern in discovering that future sci-
entists often lost a parent. Another of Galton’s discoveries is that leading
scientists were usually trained by other leading scientists and that they regard
171GRADUATE EDUCATION

that experience as critical to their own scientific development. This pattern
has been repeatedly confirmed (Roe 1953, Merton 1968, Zuckerman 1977).
Ann Roe (1953) examined the experience of eminent American scientists
from physics, biology, and the social sciences. There are many interesting
aspects to that study, but I mention only the brief autobiographical sketches of
family life and education given by each scientist. When reading these
sketches, one recognizes a most interesting omission. Not one scientist com-
mented on the type or quality of education they received, except for those
who dismissed their undergraduate days as boring. However, all vividly
remembered one scientist or teacher who had a strong impact on them and
essentially revealed the meaning of science for them. Often, contact with this
mentor was what started the eminent scientists on their successful careers.
Graduate education works because it is one of the last bastions of the
apprentice system. The student learns by working with a master, and great
masters tend to produce great students. If this hypothesis about eminent sci-
entists is valid, the availability of at least one great teacher is critical to an
effective education in science. We need teachers who inspire students so they
will achieve this potential.
Many great scientists fit this model. Claude Bernard was diverted to a
career in experimental medicine when François Magendie recognized his tal-
ent and chose Bernard to be his assistant. Antoine Lavoisier studied geology
under Étienne Cruetard, a founder of the science, and chemistry with Guil-
laume Rouelle, famous throughout Europe for his classification of salts based
on crystal structure. Johannes Kepler was started on his career by Michael
Mastlin, a famous mathematician and astronomer. Darwin walked with
Henslow. I have yet to find a great scientist who did not trace his beginning to
another great scientist. The closest I have come to an exception was Carl Lin-
naeus whose inspiration came from Aristotle through a set of Aristotle’s col-
lected works given to him by his father on his sixth birthday. In ecology, we
need only look at the success of Hutchinson’s school at Yale (Edmondson
1971): their experience with that great master gave almost all of his students
such ability and confidence that they were to define the science of ecology for
a generation or more.
The phenomenon is well described by Helmholtz whose mentor, the great
Johannes Müller, had influenced and inspired him to study physiology:
When one comes in contact with a man of first rank, the entire scale of
one’s intellectual conception is modified for life; contact with such a man
is perhaps the most interesting thing which life may have to offer.
Wise choices in graduate education. Graduate school provides both the
credentials necessary for a life in science and the training needed to use those
172 AN EDUCATION IN SCIENCE: PRESCRIPTIONS

credentials. Once touched by the peculiar passion that drives scientists, one
must go to graduate school. Selecting a school is an important step.
Since I am convinced that the most important element in the university is
the great scientist who is an inspiring teacher, this must be a major determi-
nant in choosing a graduate school. A single such teacher can make up for
years of mediocre undergraduate instruction. And in the apprenticeship of
graduate education, the presence of a great teacher becomes critical. Some
fortunate students may have found this great teacher during their undergrad-
uate years, so their choice should be clear. The others should strive to enter
the graduate environment which is most likely to provide contacts with great
scientists. One must try to work with the very best.
Although the very best masters are more likely to be found at the greatest
institutions, the right one may be found at any university regardless of size.
Institutional fame alone is a poor guide to graduate studies, because every
university has its strengths and weaknesses, and because past glories may not
signal present competence. One would do better to seek active laboratories,
because there is ample evidence that the best labs train disproportionately
more of the future leaders of the science (Zuckerman 1977).
One may also select among institutions and laboratories to find the intel-
lectual leaders within one’s chosen field, but the choice of teacher is more
crucial than the choice of material. Thus if one must chose between a jour-
neyman within one’s precise field of interest and a master in some ancillary
field, the latter is the wiser choice. Great scientists can train people to answer
more than one question, but weak teachers may not even do that.
The scientist who influences the novitiate need not be the head of the lab-
oratory. Indeed, the recognition that great scientists receive often proves to be
a heavy burden, leaving them little time to interact with their associates.
Much of the training in such laboratories may be done by younger researchers
who are themselves destined for future greatness (Zuckerman 1977). Never-
theless, because great scientists often attract, train and work with other
powerful scientists, larger laboratories and working groups are likely better
choices than smaller isolated ones.
Finding the right laboratory is not easy. Since several applicants may com-
pete for the best positions, the student should prepare for that competition
throughout the undergraduate years: by learning about the field, by earning
marks, and by winning fellowships. When the time comes to select a venue
for graduate study, one might begin with a preliminary list of potential
mentors, compiled from suggestions by instructors and fellow students, from
readings and from lectures. Such a list should emphasize the approach of the
prospective supervisor, not the material, because the approach captures the
essence of the laboratory’s science and usually endures longer. Since past sci-
173GRADUATE EDUCATION

entific fame may not ensure adequate future mentorship, the student should
research potential choices on the list thoroughly: read more of the labora-
tory’s published work, correspond with possible supervisors, and discuss the
possibilities with appropriate teachers in the undergraduate program. If pos-
sible, the prospective student should visit the lab, talk both with the potential
supervisors and with their associates, tell the supervisor what is attractive in
the laboratory’s programme, try to determine if the potential supervisor pro-
vides a positive experience for his or her present associates, and find out if
past associates have built on their experience in the lab to develop successful
independent careers.
Researching potential supervisors takes more forethought, time and effort
than most students are willing to commit. Indeed most are caught flat-footed
by any number of deadlines. As a result, prepared students often stand out
from the crowd, but it is a crowd that only the potential supervisor can see.
Since the student cannot gauge the quality of competition, the worst thing the
student can do is to decide against applying to the best laboratories because
he or she feels unworthy. Science requires confidence.
There is now a tendency to hurry the student through graduate school, sup-
posedly to increase the efficiency of education. I have little enthusiasm for
that approach, because it cheats the student of the leisure needed to develop
into an independent scientist. For similar reasons, prospective graduate stu-
dents should not settle too quickly on a topic because the first topic may not
allow exciting discoveries. Worse, it may immerse the student in stultifying
drudgery. However, the student should not take this freedom as licence for
sloth. Science is a demanding discipline and most scientists should expect to
work 60 to 100 hours a week as they develop their scientific capacities, and
for the rest of their lives. In compensation, the work is fun.
The student should try several avenues to isolate a research topic: thought-
fully considering one’s own interests, perusing current “hot topics”, searching
for an original and valid approach, and using the expertise of the supervisor
to vet and sort alternatives. In most cases, the supervisor will play a major
role in selecting an appropriate topic, because such a selection normally
requires more information and expertise than a beginner is likely to have.
Eventually, a good student will surpass the mentor in that area of specializa-
tion, thereby wresting the problem away from the master and making it that
of the student. This is an essential step because it shows that the student has
the confidence required for scientific independence.
Finally, the student will leave the privileged atmosphere of graduate
school and, usually after the respite of a post-doctoral fellowship, look for a
job. A good job in science is the best guarantee available that one will pre-
serve the freedom to express one’s scientific ideas and skills. Such jobs have
174 AN EDUCATION IN SCIENCE: PRESCRIPTIONS

always been scarce. To get one, the student must, of course, be well-trained
and do a piece of good research. The work must also be eye-catching, in the
sense that other scientists perceive the work as contributing to an important
problem. It is for this reason that I suggested the student look to hot topics.
Ideally, the work should also use a novel methodology or approach because
scientists appreciate such originality, but it must not be too new because
some members of selection committees are likely too dull to appreciate real
novelty.
Administrative Advice
To excel, a university must attract and keep people who can inspire a thirst
for science among the students. In Canada, our record for attracting and keep-
ing scientists of calibre is not good. Thomas Henry Huxley applied to the
University of Toronto for a position in zoology, but despite letters of refer-
ence from Charles Darwin and other international scientists, Huxley was
rejected in favour of a local boy. The successful candidate, who happened to
be related to the provincial prime minister, was and remains totally unknown.
Leopold Infeld, a collaborator of Albert Einstein, came to the University of
Toronto for a while, but the university chose neither to build an institute for
theoretical physics around him, nor to defend him when the local press
attacked him as too leftist; he returned to his native Poland (Infeld 1978). Sir
William Osler, who is still famous as an inspiring teacher, left McGill for
Johns Hopkins and ultimately for Britain. McGill has graduated four Nobel
laureates in science since World War II, but all achieved their eminence in the
United States. Our one laureate, Sir Ernest Rutherford, was attracted to Mon-
treal from New Zealand, and eventually left for greener pastures in England.
The brain-drain is not a particularly Canadian problem. Many smaller
countries and institutions are similarly ineffective at retaining intellectual
leaders. Why do we not attract great educators? And why can we not keep
them when we have them? Perhaps we small players neither recognize nor
want greatness. Many of our most promising individuals emigrate to places
where their ambitions can find more scope. And if they had decided to stay
home, we would interpret this as an indication of lesser stature. Since we do
not recognize greatness, we strive for an egalitarian treatment of scientists
and institutions which further insures that no individual or institution will rise
above the others. We live in a self-imposed mediocrity.
The inability of smaller institutions to hold leaders is worsened by the
advantage of great institutions in attracting such people. Merton (1968) has
discussed this phenomenon as a part of what he called “the Matthew effect in
175ADMINISTRATIVE ADVICE

Science”. In the new testament, Saint Matthew wrote, “To him who hath it
shall be given in abundance: from him who hath not it shall be taken
away.” Great institutions attract great scholars, and so grow at the expense of
smaller, less prestigious ones. Moreover, because these great scientists may
inspire one another, the advantage of the great institution continues to grow,
maintaining its preeminence and increasing its dominance.
If this is true, what can we do? I see only one answer. We must become
more elitist. We can never hope to have great scientists in all departments at
all universities. Let us therefore designate one or a few universities to become
the Harvards or Oxfords of Canada, Sweden, Italy and Australia, or of Ohio,
Ontario and Sicily. If we cannot designate whole universities as centres of
excellence, then within universities, we might specify faculties or groups of
departments for special treatment. If this is not possible, then let departments
favour particular research and teaching groups, like limnologists, behav-
iourists, forest ecologists, and neuro-biologists. If even this is not possible, let
us at least select the very best individuals among our scientific applicants.
Too often, we select against the best scientist on the grounds that the teaching
program or sub-disciplinary balance demands a different specialist, or that the
best scientist may not be the most collegial colleague. This choice ensures
mediocrity by sacrificing individual brilliance for the sake of academic
bureaucrats who treasure the equanimity of undergraduate programs and
departmental mix. Equanimity is almost always the antithesis of excellence.
An elitist choice would signal that we have accepted that we cannot teach
all things to all students, and that the value of a university education does not
depend on the number of facts or the breadth of coverage, but on the appreci-
ation of ideas. In science, ideas are theories. Great theories, like other great
ideas, are produced by inspired and dedicated individuals of great creative
vision. If we wish to promote scientific education, we must encourage those
few who can create powerful theories and, by example, train others to do so
in the master-apprentice relationship. Our first priority must be to attract
great individual scientists.
Once we have identified our favourites among universities, faculties,
departments, research groups, and individuals, our next priority is to pour
enough resources into them that we can keep the very best scientists in some
small number of disciplines. We would then have somewhere to send the very
best students — a place where they would have a good chance of encounter-
ing the odd brilliant scientist who might send them on that magical leap from
the world of pedestrian research to great science.
176 AN EDUCATION IN SCIENCE: PRESCRIPTIONS

XII The Questions of Relevance
“The English ... surpass all other nations in snobbish-
ness; our fastidious distaste for the applied sciences
... has played a large part in bringing England to the
position she occupies in the world today.”
Sir Peter Medawar
[The Limits of Science (1984)]
Previous chapters argue that contemplation of science should lead ecological
researchers, and others, to profound changes in the way we do science. If that
happened, we would be obliged to make consistent changes in other areas as
well. For example the previous chapter discussed the possibility of changing
the ways professors teach and students learn. This chapter instead focuses on
the face which science turns to the public. That too must change, if we are to
be honest with our public patrons and so keep their goodwill and support.
What Use is Science to Society?
Society has elected to support science. Politicians and bureaucrats think
research is important. Every year, each industrialized country spends millions
of dollars to pamper its scientists and researchers and to allow them to per-
form as well as they are able. There are several good reasons that this is so.
First, I can agree with Jacques Barzun (1964) that science is indeed a glo-
rious entertainment. It is a tremendous substitute for material consumption to
get us as happily as possible over that uncomfortable period between the cra-
dle and the grave. Moreover, because science is such good entertainment, it
completely occupies a whole class of technically competent, informed, think-
ing people who might otherwise make trouble because they tend to be
socially difficult. In return, society gets some good from the expense in the
vicarious pride that the country produces and supports good scientists, as well
as good writers, singers and athletes.
The entertainment value of science cannot fully explain the generosity of
society. After all, the arts, humanities, and religion offer many of the same
advantages and somewhat more entertainment, yet they enjoy far less sup-

port. Society expects more from its scientists. It expects us to be useful, and
we scientists encourage society to think so. To show that science merits sup-
port, we must show how useful it has proven to be.
There is really only one way that science is of special benefit to society. It
generates theories that let us make predictions and so look into the unknown.
These predictions help us choose courses of action that will lead to the results
society desires. Science thus helps to achieve societal objectives, but it does
not identify or judge these objectives. Many scientists believe they have the
additional duty to direct society, but that duty should be shared among all cit-
izens. Societal directions are set by policy makers who should represent the
whole of society. The only distinguishing characteristic of scientists is their
ability to produce theory.
Much of society is ignorant about the appropriate uses of science, and sci-
entists too rarely recognise any limitations to those uses. Together, scientists
and society have confounded one another with unrealistic claims and expec-
tations. Their mutual uncertainty would be reduced if both sides were more
aware of what science is, of how it differs from applied research and technol-
ogy, and of what it can hope to achieve. This chapter shows that our jobs as
biologists and ecologists would be much easier if we made explicit use of
theory to define our tasks and capacities as scientists. It also suggests that the
utility of ecology is harder to defend because we have not consistently made
use of theory in the past.
Does science differ from applied research and technology? Science has
the job of building and testing theories. For example, Einstein was definitely
acting as a scientist when he developed the relation that
Energy = Mass × Velocity
2
(25)
A person engaged in science, a scientist, need not contribute directly to the
solution of practical problems. The best the scientist can do in this respect is
to become interested in a body of data about which society is concerned. For
example, an ecologist concerned with the cycling of materials in ecosystems
might elect to study the cycling of PCB, rather than that of sodium or carbon,
because society is concerned about toxic materials in the environment. This
selection of subject matter is not applied research, but merely a diversion of
theoretical interests towards phenomena about which the society is con-
cerned.
Applied research differs from science in that its purpose is not to build or
to test theories, but to exploit potentially useful predictions from existing
theories. The person who deduces from Eq. (25) that any transformation of
matter resulting in a loss of mass would release an enormous amount of
energy, and then searches for such a transformation in the hope of exploiting
178 THE QUESTIONS OF RELEVANCE

the energy released, is an applied researcher. It is this hope, the motivation of
the research, which distinguishes applied from fundamental research.
Technology is the process of putting the results of applied research into
practice as efficiently as possible. To follow my example, once applied
researchers had identified uranium as having the best potential for the trans-
formation of mass into energy and had investigated the general conditions
under which energy release occurs, the technologists took over. They worked
out effective methods of controlling the rate of transformation and of packag-
ing the whole so it could be delivered to an appropriate site. Technology built
the atomic bomb, and got a man on the moon.
The same person can do applied research, pure science and technology,
but at different times. For example, an applied researcher who fails to achieve
the desired ends may suspect that the fault lies with theory, and begin to test
the theory whose predictions were to be exploited. To maintain my distinction
between applied research and science, I claim that this applied researcher has
become a pure scientist. The distinction is necessary to avoid working at
cross-purposes.
A facile objection to this separation is that because one worker can do all
three jobs, there is no reason to distinguish them. I disagree. To hold that no
distinction exists between these activities is like saying that because one can
eat at one time and talk at another, there is no reason to distinguish the pro-
cess of eating from the process of talking. Although eating and talking are
manifestations of a more general process called life, we still find it useful to
distinguish between them. Similarly we can categorize the activities that con-
stitute research, although we may not be able to categorize the individuals
who do them.
A more difficult objection is the problem of what to call an activity that
could be science, or applied research, or technology. For example, an individ-
ual might experiment with the fission of plutonium, not because he or she
cared about bombs or domestic energy supply, but to test the theory repre-
sented in Eq. (25). In that case, the activities that were applied research or
technology in my previous examples would be science. This might seem a
reason to reject the categorization. I choose not to do so because the distinc-
tion helps focus attention on a most important aspect of research that is often
ignored: the motivation for doing it.
Too much research is done for the same reason that a mountain is climbed
(“because it is there”), and too little time is spent questioning the motives for
doing so. We can ask why rational people have chosen to do something, and
we should expect a cogent, meaningful answer. We need to know what goals
are proposed for any particular piece of research so that we can try to deter-
mine the significance of the project and its likelihood of success. This knowl-
179WHAT USE IS SCIENCE TO SOCIETY?

edge would both help direct our research and protect us from the unrealistic
demands of society, the overweening claims by scientists, and the ineffec-
tiveness of undirected research.
Does Science Merit Support?
Up to this point, I have introduced three simple topics that receive too lit-
tle attention from university scientists: the uses of science, the predictive
aspect of theory and the differences among science, applied research and
technology. I have done so because I feel we could be more effective as teach-
ers and scientists. We can improve, but I also believe that we are unlikely to
do so unless we give serious thought to the questions that serve as headings in
this chapter.
My desire to improve is only partly altruistic. Society’s attitude to the uni-
versity scientist is changing. Our image has become tarnished and as a result,
it is likely that our working conditions will become more austere. We could
react to this by screaming at our tormenters, or we could become more useful
and effective, thus earning a better image. I prefer the latter course.
How can we evaluate our science? Obviously any group of professionals
likes to believe that its members are highly competent and without fault. Con-
sequently, I must present evidence for my unflattering opinion of university
science. I will not address the whole field but only a small part, biological
limnology and, by extension, ecology. I will look at our strengths and weak-
nesses, and speculate on procedures we might adopt to make ourselves more
effective.
Many of my views reflect personal experience with limnology in Canada,
and could be dismissed as the major failings of a minor science in a minor
country. To escape that fate, I must establish the relevance of the sample.
When we consider the size of the Canadian population and the relative new-
ness of science in Canada, the contribution of Canadian aquatic ecologists
has been remarkable. For example, data for 1982 to 1992 compiled by the
Institute for Scientific Information of Philadelphia ranks Canada second in
number of citable publications and in number of citations in both aquatic
sciences and ecology/environment. At the individual level, it is very reassur-
ing and comforting to be a Canadian limnologist abroad. Almost everyone
you meet expresses an interest in visiting Canadian limnological laboratories,
and asks about the recent work of several colleagues. Canadian limnology is
widely recognized and respected.
This reputation is well deserved. We owe it, in part, to the classic, great
limnologists of the past such as A. Huntsman, J. R. Dymond, Ronald Hayes,
180 THE QUESTIONS OF RELEVANCE

Donald Rawson, Bill Ricker, Fred Fry and so on, as well as to imports, like
Noel Hynes, Richard Vollenweider, and Dave Schindler. We can also be
proud of the great federal laboratories, The Freshwater Institute and The
Canada Centre for Inland Waters, that are in large part responsible for
keeping our reputation alive. All in all, I am very proud to be a Canadian
limnologist.
Although Canadians have reason to be pleased with our national accom-
plishments in limnology, we have no cause for smugness. Neither Canadian
limnologists nor those of other nations nor ecologists in general have reason
to be pleased when we evaluate our disciplines in relation to others.
Scientists are normally very careful not to evaluate their science relative to
other sciences. Such evaluations are difficult to do, and potentially embar-
rassing. One tool in such comparisons is the analysis of patterns of citation
because citation is one of the few quantitative measures of the impact of a
given field, an individual or a publication. Although interpretations of these
measures are disputed (Garfield 1985, Taubes 1993), I will begin with them,
because I am a quantitative scientist and because alternative indices are even
worse.
An assessment of the scientists. I began to evaluate limnology quite by
chance while leafing through Current Contents, in which Eugene Garfield
(1977a, b, c) had published a series of articles on the 250 most cited scien-
tists. To hit the top of the pops, a scientist then had to receive at least 266 cita-
tions per year (this threshold would now be considerably higher). I was curi-
ous about where Evelyn Hutchinson would appear in this list so I looked him
up. To my horror, I discovered he did not rate! Thinking that I might have
chosen my top limnologist badly, I began to look for others — Thienemann,
Naumann, Birge, Welch and so on. When I again failed, I went through the
entire list looking for an ecologist. Not one had made it. A subsequent rank-
ing for 1984 (Garfield 1986) does include a few ecologists, like MacArthur,
May, and Schoener (McIntosh 1989), but no limnologists. What does this
mean? What does it tell us about ecology in general and limnology in
particular?
First I want to rule out the comforting interpretation that our peer
group is too small to produce enough citations. The International Society
of Limnology now has about 3000 members; at the time of Garfield’s
1977 analysis, the figure was about 2500. If we consider terrestrial and
marine ecologists, we will find at least as many more. Therefore, the
minimum size of our group is 5000 scientists. If each member publishes
only one paper every second year and if each paper has (on average) 20
citations, ecologists would have collectively generated 50 000 citations per
year. In fact, there are more members, the members are more productive,
181DOES SCIENCE MERIT SUPPORT?

and reference lists are larger (Peters 1989). We therefore generate quite
enough citations to put a few ecologists on the first listing of most-cited
scientists. We can rule out the hypothesis that our group is too small to
produce superstars.
This argument does not falsify the hypotheses that large societies generate
more citations or that their leaders are likely to score higher in such compar-
isons. In other words, the presence of one or more leading practitioners
among Garfield’s immortals is no proof that other sciences are strong. They
undoubtedly share shortcomings with ecology, but that does not excuse our
showing.
At the other extreme, I reject the hypothesis that ecology attracts less
capable scientists and consequently that no ecologist is smart enough to
become a superstar (I know some colleagues in cell and molecular biology
who subscribe to this view). Many limnologists chose their field because they
were interested in and good at physics, chemistry and biology, not because
they were too stupid to become physicists or chemists. They found, in lim-
nology, a discipline that would allow them to pursue their trans-disciplinary
interests.
I have a third hypothesis. My hypothesis is that the state of our discipline
makes it impossible for limnology or ecology in general to have superstars.
The problem with ecology is that it is diffuse. It lacks focus. It isn’t going
anywhere. Ecology lacks the paradigmatic theories that identify exceptional
scientists and their exceptional work.
An assessment of the science. Perhaps some readers are still in doubt about
my meaning. If so, I will try to explain myself further by examining the char-
acteristics of a superstar scientific paper. This information is available to us
because Garfield (1977c) also listed the most highly cited papers written by
each of his superstar scientists. When I looked over this list of 250 papers, I
saw that they could be categorized into three main types: papers that describe
new theories of moderate generality, papers that describe unexpected phe-
nomena, and “methods” papers.
A new theory of moderate generality is a theory that makes very clear
predictions about a moderately large body of phenomena. Darwin’s theory
would not do. It is too general, but a less general theory, like the theory of
particulate inheritance, might count. These theories of intermediate gener-
ality help to link more general theories and concepts to theories of lower
generality and to facts. Such theories suggest many new experimental tests
or possible applications. All papers arising from these studies refer to the
paper that described the new theory of moderate generality. One reason
ecology lacks highly cited papers is that large numbers of ecologists do not
agree about what construct of great generality needs to be linked to obser-
182 THE QUESTIONS OF RELEVANCE

vation. Thus there is no place for exciting theories of moderate generality
in ecology.
New unexpected phenomena excite great interest for one of two reasons.
The first is that these observations are inconsistent with a widely accepted
theory. If this is the case, many scientists immediately attempt to falsify the
observations. If this fails, they will attempt to repeat the anomalous observa-
tions and to reconcile facts with theory. All of this work can generate many
papers that refer back to the first discovery of the anomaly.
A second situation in which new unexpected phenomena are important is
the situation that Thomas Kuhn (1962, 1970) described as a crisis. In science,
a crisis exists when a paradigm has ceased to generate soluble problems.
When this happens, leaders in the field converge on certain categories of
anomalous observations. The paper that first directed them to these observa-
tions would be highly cited. The important common feature of both situations
in which new phenomena are interesting is the existence of a generally
accepted theory to identify the norm. The lack of such norms makes it hard to
identify surprising observations in ecology.
The “methods” paper needs no definition. Everyone knows that the most
highly cited papers are methods papers. One of the best known is the 1951
paper by Lowry and co-workers on protein measurement with the folin
phenol reagent. This paper has been cited over 100000 times (to put this in
perspective, the average paper is cited about 6 times). Fewer scientists
recognize that such success is rare, that most methods papers are rejected
or uncited, and that this is especially so in ecology (Jumars 1987). Good
methods papers are hard to do.
The fact that ecologists do not produce famous methods papers is indica-
tive. God knows we have as many methodological problems as any other
group of scientists. For example, environmental toxicology has been held up
for years by our inability to analyze toxicants accurately. In fact, we still lack
acceptable methods for some commonly measured substances, like chloro-
phyll a or nitrogen. Turning the picture around, we discover that many of the
methods we use have been developed by biochemists or chemists. For exam-
ple, all of our popular methods for phosphorus analyses were developed by
others and adopted by us. Why have no ecologists developed famous
methods?
I think the answer is simple. It is very difficult to persist until a new
method is perfected. Because thorough testing takes a great deal of time and
effort, a good methods paper needs strong motivation. A large part of this
motivation is provided by the recognition that the method is absolutely essen-
tial for scientific progress and the knowledge that one’s peers need and will
use the method. In short, when the whole community of scientists recognizes
183DOES SCIENCE MERIT SUPPORT?

that certain measurements must be made, we know the methods are impor-
tant, and we are willing to make the commitment needed to produce a strong
paper about them. This recognition of important methods presupposes the
direction provided by an overriding theory.
Thus in all three classes that I could identify in Garfield’s list of famous
works, writers and users were directed by a theory that identified what is
important. I hypothesize that ecologists and limnologists do not agree about
their paradigmatic theories and therefore have failed to produce famous
papers and highly cited scientists.
Age of citation. We can test this hypothesis further by analyzing the age of
material cited in different disciplines. If a field is flagging, old papers can be
as valuable as new, so their appearance in the literature may be unheralded but
their lifespan should be long. These comparisons should make some
allowance for that spurious modernity which makes us cite the latest redis-
covery of the wheel rather than the scientist who first did the work.
Where progress in science is rapid, worthwhile papers are discovered
and used shortly after they are published. In such a discipline, papers also
go out of date very rapidly. As an example, Fig. 34 compares citation pat-
terns in ecology and biochemistry. It shows that biochemists cite propor-
184 THE QUESTIONS OF RELEVANCE
Fig. 34. The effects of age of papers, in all journals and in just ecological or biochemi-
cal journals, on the rates of citation to those papers

tionately more new papers, so the number of citations per year rises rapidly
to peak at about 3 years, and then falls off. In ecology, the initial response
is slower and less dramatic; citations rise to a low peak at about 5 years, and
then relatively slowly decline. Comparisons among other journals yield sim-
ilar data (Peters 1989, 1991a). This pattern suggests that ecologists do not
immediately recognize exciting papers as important, or are not writing such
papers.
Extent of application. In the past, some colleagues have thought this com-
parison of ecology and biochemistry unfair. They say that biochemistry dif-
fers from ecology because it has a dependent applied science, medicine. This
additional audience for biochemistry leads to more rapid application and high
citation rates. This objection actually gives additional support to my argu-
ment. Society needs an applied science of environmental management. The
reason it does not have one is that we have failed to build the foundation of
predictive theory that applied research and technology require. Therefore, I
repeat that ecology moves slowly because it lacks paradigmatic theories
which would attract citation, direct research and sustain applied research and
technology.
How can ecology merit support? I have sought to describe a situation in
science and tried to minimize the value judgements that arise from evaluation.
To the extent that I succeeded in avoiding these judgements, I have acted like
most other ecologists. We appear unconcerned with our failure to provide
theory.
I find it sad that we do not care. Society is better disposed towards ecolo-
gists than to other academic scientists. The public accepts that we need
powerful ecological science, applied ecological research and ecological
technology. Society is prepared to pay dearly for such research. Regrettably,
ecologists seem to have turned their backs on this opportunity to merit
support. At least one learned ecological society, the American Society of
Limnology and Oceanography, actually voted not to allow applied research
papers space in their journal, Limnology and Oceanography. The Ecological
Society of America has established an applied journal, Ecological Applica-
tions, but at this early stage much of the material appearing there seems as
academic as that in their established journals, Ecology and Ecological
Monographs. This has also been the fate of the British Ecological Society’s
Journal of Applied Ecology.
It seems I cannot help myself. Even when I try to change the topic, I still
end up diagnosing the ecological disease. Ecological science lacks general
theories; worse, ecologists do not recognize that deficiency or the theories of
low generality that they do have. I must complete the metaphor by suggesting
treatment, and that is how I will finish. The cure is simply to begin making
185DOES SCIENCE MERIT SUPPORT?

theories and testing predictions. The first step in this cure is to ask ourselves
questions about how successful we are as scientists:
(1) What do we want to know?
(2) What predictions do current theories make?
(3) How can we improve old theories?
(4) What research is likely to provide new theories?
I do not know how we will answer these questions. I suspect that the last
two are unanswerable because theory improvement requires creative insight.
However, I believe that the exercise of groping for answers will help give
ecologists the directions we presently lack. We will then be in a position to
earn the support of society, to return our patrons’ investment many times
over, and to fulfil our moral obligations to humanity and to nature.
186 THE QUESTIONS OF RELEVANCE

XIII Funding Decisions
“The course I propose for the discovery of sci-
ences is such as leaves but little to the acute-
ness and strengths of wits, but places all wits
and understandings nearly on a level.”
Francis Bacon
[Novum Organum (1621)]
“Quite ordinary people can be good at science.”
Sir Peter Medawar
[The Limits of Science (1984)]
The search for funds, the writing of proposals and the evaluation of grant
requests by our peers are major elements in the lives of most contemporary
scientists. A clear grasp of what science is and what scientists can do is an
indispensable aid in all those processes. This chapter therefore applies
lessons from my contemplation of science to the various facets of grantsman-
ship. It identifies the legitimate expectations one can have of research pro-
posals, discusses the gamble involved in trying to fund scientific creativity,
and warns against some current practices in science. The chapter ends with an
extended analysis of multi-disciplinary team research as an example of how
the granting agencies’ misconceptions about science can make science more
difficult.
The Central Problem for Research Funding
Basically, a grant proposal promises to do a certain set of tasks in return
for a specified sum of money. The problem for the proponent is to describe
future research so that its promise is great. For the panel, the problem is to
determine which of all the promises it reads are most likely to succeed.
One of the first discoveries one makes as a part of peer review is that ecol-
ogists have been largely untouched by the last three hundred years of philos-
ophy: philosophers showed long ago that the applicants cannot possibly keep

many of the promises their applications make. We all need a better grasp of
what research can and cannot do.
Some applicants imply that theories can be produced when required, like
rabbits from a magician’s hat. They promise that if given enough funds, they
will make predictions about phenomena of immediate interest to society and
resolve long-standing questions in science. The continual barrage of promises
raises unjustifiable expectations in the public, administrators and legislators
who need useful answers from scientists.
Other applicants seem to subscribe to the old belief that theories are pro-
duced by a logical process based on data collection, that theories can be ver-
ified by further observations, and that after sufficient verification, they will be
accepted as true natural laws. This set of beliefs assumes that if we make the
correct observations carefully and thoroughly, the facts will logically and
inevitably lead us to discover true theory. Consequently, applicants believe
that if they can promise to make new observations correctly, then they will
also be able to derive, from these observations, a new theory that would gen-
erate predictions needed by society. David Hume, building on the earlier
work of Locke, Berkeley and others, showed in the middle of the 18th century
that this ancient belief is false.
We now know (to the extent that we can know anything) that a body of
specific facts can never lead logically to more general statements, that logic
cannot lead us to discover a new theory. Medawar (1984) called this idea the
law of conservation of information: “No process of logical reasoning — no
mere act of mind or computer-programmable operation can enlarge the infor-
mation content of the axioms and premises or observations from which it
proceeds.” Instead, any body of facts can give rise to an infinite series of dif-
ferent theories (Chapter I). The competing theories must subsequently be
winnowed and selected by scientific tests.
Scientific theories are a product of an inspired human mind via a process
we neither understand nor control. We cannot promise to be inspired, and for
that reason, scientists cannot design a project that is sure to produce a theory.
Since no one can promise to be inspired, grant reviewers must be suspicious
of any proposal built around an attempt to create a theory. Yet any granting
panel finds application after application that proposes to do just this. Some-
times these proposals even merit funding.
Reasonable expectations from research. Scientists cannot promise to
create a theory, but they can undertake other aspects of research where the
product is fairly certain. Most of these are premised on the prior existence of
some theory, because scientists use accepted theories as paradigms to direct
their research effort. In the absence of theory, ecologists lack direction to do
anything, and research flounders. For example, I believe that the current
188 FUNDING DECISIONS

drifting of ecology reflects the non-existence of the theories ecologists want
and their unwillingness to recognize the theories they have. The goal of this
section is to show how theory structures research and research proposals,
and its motive is to further stress the need for theory in everything scientists
do.
Theory testing. If a theory is any good, it specifies certain observations as
more likely to be observed under given conditions, and others as improbable.
Our grant proposals can promise to test these predictions. A scientist can
make this promise because the test merely involves gathering data specified
in the theory. In this case, the reviewer can expect a clear exposition of the
theory, the logical derivation of deductions from that theory, a statement of
the observations that will be made, specification of what observations would
be inconsistent with the theory, a description of the methods that will be used
to make them, and a statement of the accuracy with which they will be made.
If one or more of the specified, improbable events occurs in a series of tests,
we are likely to reject the theory. If a predicted event occurs, our faith in that
theory becomes stronger than our faith in competing theories. Theory testing
is one scientific activity from which we can expect results.
Since I have repeatedly charged that ecological theories are flawed, unpre-
dictive, unrecognized or non-existent, theory testing may seem irrelevant to
ecology. This need not be so. All that is required is that the proponents elab-
orate the theory themselves before approaching the granting agency. In other
words, we must think about what we are going to do before we do it. This is
not a new paradigm for ecological research, because theory creation has
always been an individual effort, but it does require that the proponent
describe the theoretical prototype and render it interesting to the granting
panel. It also requires that the panel and agency recognize the importance of
funding research that is based on theory, even when the community cannot
agree which theories constitute its paradigms.
Applied research. The preceding chapter defined applied research as a
search for the potentially useful predictions of existing scientific theories.
Scientists can justifiably propose to do an applied research project because, as
I have defined it, applied research does not commit us to the production of
new theory. Instead applied research tests existing theory within the limited
framework of application to socially interesting problems. Neither are we
committed to succeed if the existing theory is wrong.
Technology. Chapter XII also defined technology as the process of extract-
ing maximal benefits from the successes of applied research. This definition
does not preclude the possibility of technological advance on its own nor does
it deny that at times science has developed in the wake of technological
advance. However, I am concerned with designing projects in ecology and in
189THE CENTRAL PROBLEM FOR RESEARCH FUNDING

that field, I believe technological projects will largely exploit the results of
applied research. This again is suitable material for a project, because we do
not have to invent new theories to succeed in technology. We merely have to
find the best ways of applying those that already exist.
Surveillance monitoring. Monitoring involves the routine measurement
of a set of predetermined variables. In surveillance monitoring, the variables
to be measured are selected because they give a sensitive measure of the
state of the system at minimum cost. This process of assessment should not
be confused with research monitoring whose purpose is to discover relation-
ships among the monitored variables, and so to develop new patterns and
generalities.
Programs of surveillance monitoring require theory, but are not intended
to test those theories. They involve simple fact-gathering and can therefore
promise to deliver specified results (tables of measured values) within a spec-
ified time. These facts are empty by themselves, but the collection may be
useful, because these facts may be used as “predictors” or “independent vari-
ables” in existing theories.
For example, mussel-watch programmes to detect phytotoxins in commer-
cial mussels (Mytilus edulis) are a form of surveillance monitoring. In
Canada, when ambient levels of toxin in commercial mussels reach the criti-
cal value of 20 µg l
–1
, potentially contaminated shellfish are kept from market
(Novaczek et al. 1992). Surveillance monitoring is needed because no
accepted theory predicts phytotoxin concentration. The programme is still
derived from theory, because only a theory can associate a given toxin level in
shellfish with the poisoning of human consumers. Nevertheless, the monitor-
ing programme is a form of technology not a test of theory, because we do not
wait to see if the poisoning would actually occur. In other words, the theory is
not tested, it simply directs sampling.
Surveillance monitoring is normally undertaken by consulting firms or
government agencies, not by scientists. If possible, a programme monitors
“primary variables”, those that are of greatest interest to the society. For
example, in eutrophication monitoring, primary variables may be greenness
of the water, fish yield, water taste and odour. Since the primary variables are
not always easy to measure — greenness varies in space and time, fish catch
is influenced by human whims and accidents, taste and odour are hard to
quantify — many monitoring programmes focus on “secondary variables”.
Secondary variables are related to the primary variables by theory, but are
more easily measured (“simplifying secondary variables”: e.g. Secchi disc
depth), vary less (“integrating secondary variables”: e.g. total phosphorus
concentration, hypolimnetic oxygen content) or warn us that the primary
variables that characterize the system are likely to change (“causal secondary
190 FUNDING DECISIONS

variables”: e.g. the number of permits issued for logging or building in the
catchment). There is also place for “confounding variables”, like temperature
or solar radiation, that might obscure trends in the primary and secondary
variables.
Monitoring programmes begin with a crucial decision about what should
be measured, and any proposal for surveillance monitoring must include a
reasoned defense of the variables selected. Since the only acceptable defense
for surveillance is that the variables function in a theory to tell us something
about the primary variables of interest, proposals for surveillance monitoring
must specify the relevant theories.
Since ecology recognizes few relevant theories, many programmes of
surveillance monitoring have not been well defined. As a result, they end in
indefensibly large collections of almost whimsically selected variables. For
example, I was once involved in developing a strategy to monitor eutrophica-
tion in the Great Lakes. As part of this process, three authoritative lists of
potential variables were developed: one from the OECD eutrophication mon-
itoring programme, one from G. Fred Lee, and one from Jack Vallentyne.
These lists (Table 18, overleaf) each identify between 9 and 27 essential vari-
ables, and together include a total of 38 characteristics. The types of variables
considered essential also differed. OECD listed a series of anions and cations,
whereas Lee and Vallentyne ignored them all. Only Lee considered the vari-
ous forms of phosphorus and nitrogen to be essential, and only Vallentyne
recommended any measurements involving benthos or sediments. One may
conclude that different authorities used different criteria to compile their lists,
but these criteria were not made explicit.
The clearest implication of the discordance among the lists in Table 18 is
that the more opinions sought, the more variables will be considered essen-
tial. Surveillance monitoring risks a thoughtless accretion of variables, and
this is especially likely if the programme is developed by a committee. The
only protection for the proponent and for the reviewer is to insist that all mon-
itored variables be tied explicitly and tightly to theory.
How to gamble with research funds. Avenues of research that confi-
dently promise scientific advance are premised on the existence of theory. For
me, such ecological research is exciting because such a programme must con-
tain a rare treasure, testable ecological theory. In contrast, research that is not
premised on theory is almost indefensible.
Because ecology seems to lack theory, but obviously abounds with impor-
tant fundamental and applied problems, scientists often propose to create new
theory to address those problems. Any such proposal is a gamble, and if the
reviewers and the agency administrators decide to fund such a proposal, they
are gambling with research funds. The question I address in this section con-
191THE CENTRAL PROBLEM FOR RESEARCH FUNDING

192 FUNDING DECISIONS
OECD Lee Vallentyne
Physical variables
Temperature
*** ***
Conductivity
*** ***
Water colour
***
Turbidity
**
Secchi disc
*** ***
Solar radiation
***
Chemical variables
pH
*** ***
Dissolved O
2
*** *** *
Total P
*** ***
Soluble P
***
Soluble reactive P
***
Total N
***
Nitrate
***
Ammonia
***
Organic N
***
SiO
2
*** ***
Alkalinity
*** ***
Acidity
***
Ca
***
Mg
***
Na
***
K
***
SO
4
***
Cl
***
Total Fe
***
H
2
S
***
Trace elements
*
Nutrient load
***
Biological variables
Chlorophyll a
*** *** ***
Particulate organic C
***
Primary production
***
Algal dominants
*** ***
Algal genera
* ***
Zooplankton counts
* ***
Benthic invertebrates
***
Sediment cores for:
Pollen
***
Diatoms
***
Invertebrates
***
Table 18. Variables thought to be essential (
***
) or desirable (
*
) measurements in a pro-
gramme to monitor eutrophication of lakes, according to three authorities in the early to
mid-1970’s

cerns the odds of the gamble. We know that a new theory cannot be guaran-
teed, but we can shorten the odds in its favour.
Empirical theory. When no brilliant insights are available, one way to
shorten the odds against theoretical success is to develop the simplest kind of
theory. These are empirical, interpolative or correlative theories, and are
based on the identification, analysis and description of naturally occurring
patterns in phenomena of interest: identify the variable of interest, describe
the distribution of its values, and perhaps, explain some of this variation sta-
tistically by reference to other variables. This has been the universal path to
theory in other sciences and is the one which granting agencies should pref-
erentially support.
If the identification of appropriate dependent or independent variables
seems a challenge, there are ways to identify likely possibilities. For example,
one can test the applicability of an existing theory beyond established
limits. We might extrapolate a regression beyond the data base on which it
was built, as Dillon and Rigler (1974a) developed a general phosphorus-
chlorophyll regression by applying Sakamoto’s (1966) relation to non-
Japanese lakes. Or we might attempt to adapt a theory built for one type of
system to another type, as Smith et al. (1984) showed that phosphorus-
chlorophyll response models developed for temperate lakes also apply in the
sub-Arctic.
Another way to increase the likelihood of success in theory-building is to
exploit possible homologies to existing theories. For example, one can be rea-
sonably confident that most physiological rates will be affected by body size
(Peters 1983, Calder 1984), so as yet uncorrelated autecological processes,
like the rate of uptake of organic contaminants or the time to die from expo-
sure to heavy metals, are likely to be predictable from body size. Because the
effects of size are so conservative (Peters 1983, Calder 1984) we can even
hypothesize that the exponents will be 0.75 for individual rates of uptake and
0.25 for physiological times. The homology can be less close: the widespread
occurrence of resource limitation suggests that successful theories to predict
biomass might well use resource level as a predictor (Peters 1991a); similarly,
Keddy (1989) posits, on the basis of a small series of comparisons, that com-
petitive relations will be most easily predicted by differences in competitor
size.
The characteristic of all these relations and patterns is that they can be
elaborated as concrete, testable theories with only a small amount of thought.
Pattern identification is sufficiently easy that a rudimentary theory can be
expected as part of the grant proposal, perhaps based on preliminary results or
literature survey. If ecologists were satisfied with simple patterns as theories,
we would recognize that our science already includes a vast corpus of theory
193THE CENTRAL PROBLEM FOR RESEARCH FUNDING

and potential theory. If we recognized these relations for what they are, we
would soon be led to theory-testing, the soundest basis for research proposals.
Research monitoring. At times, scientists may propose surveillance moni-
toring, but they are usually interested in more than simply gathering facts.
They may believe that the facts will provide inspiration for a new theory; or
they may want to make other measurements that involve little expense over
and above the cost of surveillance. Scientists often wish to collect facts that
were not previously known because they have a gut feeling those facts will be
important, yet the only way they can afford to play their hunch is under the
guise of surveillance monitoring. Such a proposal should be seriously consid-
ered, because the end result can be a more efficient use of research funds, but
the lure of economy should not obscure the fact that this type of work
promises to create theory. Research monitoring must be treated with the scep-
ticism that risky research deserves.
Explanatory theory. There are also ways to increase the odds against
creating theory. The most difficult thing a scientist can do is to develop a new
explanatory theory, and the more general the theory the harder the task.
Regrettably, the majority of ecologists would not be satisfied by seeking pat-
terns in nature. They yearn to develop general, explanatory theories, and by
trying to do so, they make their tasks vastly more difficult.
Because the demands of creating general explanatory theory are great,
such theories are exceptionally rare. The few people who succeeded are
revered as geniuses, like Newton, Einstein, Mendel and Darwin. Perhaps it is
significant that in these particular cases, no proposal was written and the
work was not publicly funded. Indeed, proposals to develop theories of high
generality rarely come to granting agencies, and when they do, they are usu-
ally not funded. The gamble is too large and the pay-off too uncertain.
Science benefits immensely from genius, but researchers of such quality
are too rare to be the basis of science policy. Heroes are not models for
the common-or-garden variety of research proposal. Administrators since
Francis Bacon (Eiseley 1973) have recognized that science policy must be
designed for the best of the common scientists. The agencies hope to recog-
nize and pamper genius if it appears, but they cannot found a policy premised
on the existence of genius. Perhaps the best we can expect is enough funding
for our leading scientists that the select few will find both the freedom and
ability to create in this heroic sense.
Some realities of ecological research. Effective research can be designed
to test an existing theory, to collect data so a theory may be applied, or to use
existing theory in applied research and technology. All of these projects are
premised on the existence of theory. We must also accept that, although no
project can be designed to produce a new theory, we must find ways to fund
194 FUNDING DECISIONS

scientists in the hope that some will succeed in creating a theory anyway.
However, much of what grant applicants propose does not fit any of these
research strategies.
Many scientists try to show how good existing theories are, and their pro-
posals are therefore designed to corroborate, not test. These researchers are
doing normal science (Kuhn 1962, 1970). Some ecological proposals adopt a
similar strategy, even in the absence of theories. They focus on a topic that
interests some sub-component of ecology (the 1991 index issue of The Amer-
ican Naturalist suggests these topics might include, among others, altruism,
sex ratios, density dependence, food web connectivity, patch selection, poly-
gyny, and philopatry) and strive to interpret associated phenomena in terms of
the concepts and theories of their sub-subdiscipline. This methodology
recalls Aristotle’s search for circles in nature. Most such work is usually con-
firmatory and therefore serves to entrench ideas in the sub-subdiscipline.
Such studies are among the least worthy of support. They strive to defend the
status quo, rather than to improve the science.
The Dream of Multi-Disciplinary Environmental Science
Granting agencies have a necessarily close relation to the scientists they
support. As a result, the proponents’ misconceptions about the nature of ecol-
ogy reappear among reviewers and agency representatives. However, because
the agencies are so powerful in shaping science, the misconceptions of their
agents can be particularly dangerous. Given that science continues to flour-
ish, such agencies more or less succeed in their mandate, but whether this
success is because of their interference or in spite of it is usually hard to
judge.
Sometimes, agency initiatives are misplaced. One example of the latter
is the decision to encourage multi-disciplinary research in environmental
science. This initiative reveals a profound failure to appreciate the nature of
science. Yet similar initiatives have been made by most agencies around the
world.
The advantages of team research. I intend to criticize the emphasis on
scientific research by multi-disciplinary teams as counter-productive in ecol-
ogy, so let me begin by stressing that I am not criticizing team research in
general, teams of researchers, or multi-disciplinary approaches to applied
research and technology.
There are many projects for which teams are essential, and even more for
which teams have proven useful and effective. Teams of researchers provide
an economy of scale; two researchers may need no more spectrophotometers,
195THE DREAM OF MULTI-DISCIPLINARY ENVIRONMENTAL SCIENCE

scintillation counters, or chromatographs than one. Sympathetic colleagues
provide support in ways ranging from helping to move the furniture to tem-
pering a personal crisis in one member of the group. Such colleagues, work-
ing on more-or-less related fields, represent informal teams and are invalu-
able as sources of ideas, criticism, and information. I believe too that we
should promote collegial teams so they become centres of excellence, not
because team research is better than individual research or because teams are
more productive than individuals (they are not; Cohen 1991), but because
teams provide a context within which individual scientists are more likely to
flourish (Chapter XI). It is appropriate to encourage research teams, provided
this encouragement does not discourage the single researchers who have
always been the source of most of the new ideas in science.
More coordinated teams are necessary when a single scientist is unable to
test the predictions of existing theories, perhaps because the tests require too
many data collected over too short an interval or too vast a scale. Such prob-
lems must be addressed by team research, but the teams need not be multi-
disciplinary. Indeed, the need for teams of workers who are interested in
addressing the same problem would seem more likely to arise in single disci-
plines with strong central theories. Such theories will attract people to test the
theory the group holds in common. Under this scenario, the requirement for
team research is generated by the science. A funding agency should have
policies in place to accommodate team research proposals as they arise, but
this is very different from the current inclination to impose team research on
sciences which are not ready for them.
Multi-disciplinary teams have repeatedly demonstrated their utility in
applied research and technology. The Manhattan project and NASA are
among the best examples. In those cases, the scientific problems were largely
solved when the team was formed, but immense challenges remained in
application and technology. The basic problem was to construct a mechanism
along established principles that could perform a single well-specified job.
Multi-disciplinary teams function well in attacking such questions. The
mechanism can be reduced to its parts, farmed out among different experts for
solution, and the many parts eventually assembled as the whole machine. The
machine is then tested to see if the assemblage works. If some parts fail, they
can be redeveloped by the appropriate teams. In summary, the success of
multi-disciplinary projects demonstrates the advantages of the guiding theo-
retical structure in mature sciences, of the greater financial efficiency and
camaraderie in teams of researchers, and of a specified goal in applied
research.
The problems of multi-disciplinary research in ecology. I think it is fair
to say that, although scientists need to be continually reminded of their short-
196 FUNDING DECISIONS

comings, some of us ecologists have been bitterly aware of ours for decades.
We have tried to remedy the problem. Our most common response was prob-
ably to ask for more money, but another common response was to form multi-
disciplinary teams to attack the problem.
Before I begin to tell my tale I must carefully define the type of ecologi-
cal question that I am addressing. There are many types of ecological re-
search, and I am discussing only one, the hardest and the most important.
When we really run into trouble and where we have had the poorest record
of success is in trying to forecast the effects of human disturbance on any
ecosystem. The question we almost always fail to answer is “What will
happen if...?” A good example of this type of question is the request for an
impact statement: What will happen to the organisms in a given stream if a
dam is built across it? or What will happen if a factory discharges its wastes
into the stream?
Ecologists hailed the multi-disciplinary approach as a panacea for such
questions. Now we have had enough experience to ask if it has lived up to
expectation. I suggest that the multi-disciplinary approach has not been as
successful as we hoped and that there are two reasons for its failure. First,
these questions ask us to predict in areas where we lack explicit theory, and
second they require that we use theories from many different scientific disci-
plines. Neither difficulty is addressed well by research teams.
Our failure to foresee environmental problems. I have said that we have
proven unsuccessful at ecological forecasting, but I should explain myself.
What I mean is that none of our major environmental problems was pre-
dicted. Instead of warning society that a problem would develop, ecologists
have always had to tell society that a problem has developed. Here are a few
examples.
(1) The eutrophication of our inland waters was documented, not pre-
dicted. Many of our lakes had already turned green at the surface and anaero-
bic at the bottom long before we decided whether phosphorus, nitrogen or
carbon was the element limiting production in lakes.
(2) We condoned the use of DDT to control insect pests without predicting
its bio-accumulation and bio-concentration, or its harmful effects on non-
target species. We discovered the danger of DDT only when it had already
driven several species towards extinction.
(3) We did not predict the accumulation of contaminants, like mercury or
PCB’s, in freshwater fishes. We discovered it only after contaminants had
concentrated to dangerous levels.
(4) While ecologists in Canada were studying the problem of acidification
near the nickel smelters at Sudbury, Ontario, just as they had a generation
before at the Trail smelter in British Columbia, acid precipitation from coal-
197THE DREAM OF MULTI-DISCIPLINARY ENVIRONMENTAL SCIENCE

fuelled generating plants all over eastern North America was quietly acidify-
ing thousands of lakes from New York and Ontario through Québec to New
Brunswick.
In each of these examples, ecologists performed a useful service to soci-
ety, because we discovered the problem. However, in every case we failed to
predict the problem before it arose. Since predictions arise from theories, we
could only forecast such problems if we had a theory of ecosystem response.
If no such theories exist, the most important task for concerned ecologists
must be the creation of pertinent theories.
The scientific activity for which team research is least appropriate is the
creation of new theory. This step depends on inspiration, and inspiration
occurs inside a single human mind. A team cannot undertake to create a new
theory. Indeed, since teams often act like committees, they may discourage
creativity and new directions in research, by striving for consensus. Until
ecology has theories that are general enough to interest many researchers,
team research in ecology will largely be premature.
A false solution. The second reason that environmental forecasts are diffi-
cult is that the required general theory will almost always derive from exist-
ing theories of lesser generality from several scientific disciplines. The gen-
eral theory can then be called multi-disciplinary. The flaw in our approach to
such problems is that we assumed that the development of such theories
required multi-disciplinary teams of uni-disciplinary individuals. The multi-
disciplinary approach to ecology failed, as I think it had to fail, because that
central assumption was erroneous.
We have very good reasons for thinking that progress in environmental
science will be slow if we work within the boundaries of the traditional disci-
plines. Environmental problems never attended university, and therefore do
not respect the conventional compartmentalization of scientific knowledge.
For example, to predict the distribution and effects of air-borne contaminants,
we probably need knowledge from chemical engineering, meteorology, atmo-
spheric physics, phase, surface and inorganic chemistry, botany, soil science,
hydrology, limnology, biochemistry, ecology and so on. Our mistake arises
when we extend this argument to scientists and argue that to solve multi-
disciplinary problems, we must assemble multi-disciplinary teams. That
response was and is counter-productive.
Since we need new theories, we must depend on the individual scientist,
and since the problems transcend conventional disciplines, we need individ-
ual scientists whose training and interests are similarly unconstrained. What
we really need to solve such problems is a new breed of scientists, single
“trans-disciplinary” creators whose breadth is such that they can cross tradi-
tional disciplinary boundaries.
198 FUNDING DECISIONS

The time-honoured solution. We have failed to produce predictive theo-
ries, and the institution of multi-disciplinary research teams has contributed
to this failure. What is the alternative? To find this alternative, and to discover
the flaw in the multi-disciplinary approach, I suggest you examine very care-
fully the history of the branch of science that you know best. When you do
this, you will discover that major scientific progress has always taken place
along a very well-defined pathway:
(1) Identify the class of systems under study.
(2) Observe the behaviour of these systems intact.
(3) Discover pattern and quantify it as empirical theory.
(4) Study the system components, probably by mechanistic dissection.
(5) Replace empirical theory with explanatory theory.
The important point for this discussion is that if successful sciences stud-
ied isolated components of their system at all, they did so only after they had
achieved a theory. Successful scientists discover the regularities of their sys-
tem before they try to explain them. Reductionist methods are used only in
step 4 where they provide inspiration for step 5.
I should digress momentarily to say that although I may appear to be
taking a hard line, I am actually middle-of-the-road. There are very strong
arguments leading to the conclusion that it is never appropriate to study iso-
lated subsystems (Peters 1991a), unless it is the behaviour of isolated sub-
systems that we want to predict. I went into these arguments in Chapter VII,
but here I have taken a weaker position that is justified on historical grounds
only.
What strategy is appropriate to ecology? Earlier I gave examples of our
inability to predict environmental problems. We discovered them after the
fact. This suggests to me that environmental science is at a very early stage. It
has not yet produced an adequate set of empirical theories. If my conclusion
is correct, we should concentrate our efforts at steps 1 and 2 in the sequence.
We should attempt to define and classify the systems about which we need
predictions. When this is done, we should study the behaviour of these sys-
tems. Perhaps then, we will produce the empirical patterns and theories
required for further scientific advance.
Where does the multi-disciplinary approach lead? With these guidelines
in mind let us now think about the multi-disciplinary team, and ask ourselves
how it will approach multi-disciplinary problems. Experience shows that the
multi-disciplinary group quickly decides to dissect the system and to study its
fragments by the techniques appropriate to each discipline. For the physicist
or chemist, this is the approach to which they have been conditioned. They
come from advanced disciplines that are replete with highly explanatory
199THE DREAM OF MULTI-DISCIPLINARY ENVIRONMENTAL SCIENCE

theories. In these disciplines, the tradition of dissecting the system into
smaller and smaller fragments is well established. They succeed to the extent
that commonly held, general theories keep the whole enterprise together.
The ecologist in the team could be expected to act differently because gen-
eral theories in ecology are almost non-existent. However, as I have argued
previously, most ecologists reject the lessons of history. They have decided
that because their systems are more complex than those studied by other sci-
ences, ecologists must reverse the normal process of science. They begin at
step 4. They are methodological reductionists, looking for an explanatory
theory before they have discovered the regularities that need to be explained.
Because the members of the multi-disciplinary team tend to reductionism
so does the team. In the extreme case, each member engages in reductionist
studies aimed at furthering development of his or her discipline rather than in
developing new theories for ecology.
This is the flaw in the multi-disciplinary approach.
Where do we go from here? Since I cannot end on this depressing note,
I will suggest very briefly that there is a viable alternative to the multi-
disciplinary approach. To develop an alternative, I think we must first accept
that ecology and limnology are disciplines in their own right. They depend on
theories of chemistry, physics and biology, but they are not merely an aggre-
gation of these. Because the ecological sciences must predict about a differ-
ent class of systems, their problems are different from the problems of the
chemist, physicist and biologist.
Ecological solutions require a trans-disciplinary generation of scientists.
We must learn to use the theories of the traditional academic disciplines as
tools and to stop making their theories the object of our research. In addition,
we must learn how to train new scientists as ecologists or environmentalists.
They must realize that they are not physicists or biologists who were not for-
tunate enough to get a job in real science. They are scientists with primary
allegiance to a different natural system. In other words, they must be trained
to see their speciality as a science in its own right, not just as an applied
appendage of some “real” science.
200 FUNDING DECISIONS

XIV Darwin and Evolutionary Science
“The amount of assumption and reasoning in a
vicious circle involved in these [current evolu-
tionary theories] renders it certain that none of
them can long survive.”
Sir William Dawson
[Modern Ideas of Evolution (1890)]
In this chapter, I want to show how an appreciation of science can help to
interpret the history of our discipline, and conversely, how a knowledge of
our history can help us to understand our science. I will begin by describing
how Darwin’s observations on the Galapagos may have allowed him to fal-
sify competing theories of the origin of the species and so to develop his own
theory based on that experience. My idea may be wrong, but I find that it
makes Darwin and the Galapagos more interesting and vital than no idea at
all. Having given Darwin some of his due as a scientist, I will then examine
some critics of Darwin’s theory to see if they are equally robust. The purpose
of the exercise is to show that a grasp of the nature of science can clarify the
context of our work.
Like other biologists, I have always admired Charles Darwin. I admire
him because he dedicated decades of his life to an articulated series of stud-
ies involving different methods and organisms, all directed to the develop-
ment of a coherent vision of the living world, because his experimental work
was carefully considered and carefully done, because he saw further than the
rest of us (Ghiselin 1969) and because he succeeded in science without aban-
doning either his family or his community. Nevertheless, I am dismayed
because he is rarely treated with the respect and dignity that a scientist of his
stature has earned.
Darwin is either deified by his followers or vilified by his foes. Both
extremes are inappropriate. Darwin was one of the greatest scientists of all
time, and his ideas should be treated accordingly. We do him greatest honour
when we treat his ideas as instances of contemporary science, open to test,
rejection and revision. If we elevate his writings to the level of sacred texts,
we do him and his science as much insult as those who reject his views as
inconsistent with a literal reading of the Bible. Science needs no dogma.

Darwin on the Galapagos
Biologists all know that while visiting this tiny archipelago over 150 years
ago, Darwin made the observations that sent him reluctantly and inevitably
along a path he had known about but previously avoided. The result of his
subsequent labours was one of the few theoretical revolutions that biology
has ever experienced. It was a revolution that changed our world view dra-
matically and irreversibly. The effects spread like a chain reaction, first
through biology by precipitating new lines of research in comparative
anatomy, comparative embryology, population genetics, etc. Subsequently,
history, economics, ethics, metaphysics, sociology and even art and poetry
were changed beyond recognition by new attitudes engendered by the theory
of evolution by natural selection. All this started on Darwin’s trip to the Gala-
pagos!
The observations Darwin made must have been remarkable. But what
were they? After asking yourself, ask some colleagues to see if they know
what observations he made. If you and your colleagues are as well informed
as I was when I first asked this question, the answers will be disappointing:
“Didn’t he observe finches with different beaks adapted to different foods?”,
or, “He saw the results of finches and turtles evolving to adapt themselves to
new environments.” Although such answers seem satisfactory on first inspec-
tion, a little thought shows us that they are not. If Darwin merely saw that the
body parts of different animals were well suited to a function and to the habits
of those animals he saw nothing new. No one in his time, or before, expected
a lion to live on grass or a cow to be a carnivore. To tell us that Darwin saw
the results of evolution through natural selection on the Galapagos is to
phrase the answer in terms that were meaningless before his theory existed
and to imply that the results of natural selection were not to be seen elsewhere
in the world.
If our standard answers are unsatisfactory, where can we go to find better
ones? Years of conditioning as students and teachers will probably send us to
introductory biology texts. Here we will find more disappointment. By and
large the texts give answers just like ours, although more detailed and accom-
panied by attractive illustrations. At least our own ignorance has been
explained. The answer to my question was unknown when the text-books
from which we learned our biology were written.
Why, then, were these answers either forgotten or undiscovered? The first
reason I suggest is that we mostly subscribe to the old-fashioned “truth theory
of science”: all current theories are true, all discarded theories are false. And
because of this we shy away from serious discussion of rejected theories.
Consequently we were not taught, are not really interested in, and do not
202 DARWIN AND EVOLUTIONARY SCIENCE

teach rejected theories or the reasons theories were rejected. Therefore the
observations that led to the rejection of a theory are not significant to us. In
this particular case however, there is another source of our difficulty and that
is that Darwin never said what observations influenced him while on the
Galapagos. All he gave were some pretty broad hints in his diary from the
voyage of the ‘Beagle’.
Because I believe the events leading up to the rejection of a theory are
important in helping us to understand our science I will try to interpret
Darwin’s hints. To begin, we must know what theory or theories existed in
Darwin’s time. The old biblical theory of creation at a single time and place
had been long abandoned, and two alternatives were competing for domi-
nance. One, proposed by Lamarck, said that God created life with a built-in
drive towards perfection. This force alone caused slow evolution, but the pro-
cess was accelerated and often diverted by the environment. Thus, similar
environments would produce similar species. The competing theory was
proposed by Georges Cuvier, and Charles Darwin encountered a modified
version of this theory in Lyell’s geology text which he read during the
voyage. This theory explained the known facts about fossils and animal dis-
tribution by postulating different centres and times of creation. It did not
allow for production of new species by slow evolutionary change.
Now let us consider the Galapagos in the light of these two conflicting
theories that formed Darwin’s world view. He saw a group of islands that
were obviously of volcanic origin and, unlike South America, obviously very
recent because the lava streams showed no signs of erosion. He saw birds and
plants that were peculiar to the islands, but similar to species he had seen in
South America. Even more surprisingly, the species were unlike anything he
had seen on the Cape Verde islands near Africa, despite the similar climates
and geologies of the two archipelagoes.
Can we explain these facts in the light of Lyell’s or Lamarck’s theo-
ries? According to Lyell, the organisms of the Galapagos, being endemic,
must have been created there. Since South America is ancient and the
Galapagos were recent, they must have been populated by two separate
acts of creation widely separated in time. If so, why would the Author of
Nature, who creates species to suit the conditions they will encounter,
create species for the Galapagos on the South American rather than the
Cape Verde plan? The facts just do not make sense in relation to the the-
ory and to our image of the Creator. Then let us follow Lamarck and see
what he would predict if the Galapagos had been populated by migrants
from South America that subsequently evolved to suit the conditions they
found on the Galapagos. This is where the finches become relevant, be-
cause Darwin had seen one, and only one, species of finch living under
203DARWIN ON THE GALAPAGOS

incredibly diverse conditions all along the west coast of South America.
Clearly this must have been the species that first colonized the Galapagos
where it gave rise to eleven different species. But if species evolved to
adapt to the conditions they encounter, why was there one species in
South America and eleven in the climatically and geologically uniform
Galapagos? Surely Lamarck’s theory would have predicted just the oppo-
site!
Now that we know the theories, we can see that the facts of the Galapagos
seemed to contradict them. Of course, the facts did not force Darwin to pos-
tulate his theory, but they did send him on a quest that led eventually to his
theory. How his thinking was led to ideas about natural selection is quite a
different story that has nothing to do with the Galapagos. In fact, because that
involved a creative step it may have no rational explanation at all, whereas the
rejection of existing theories on the evidence is simply good science by a
great scientist.
Critics of Darwin
McGill University, where I am writing these notes, is well known in evo-
lutionary circles as one of the last outposts of anti-evolutionary thinking. The
famous anti-Darwinist that gave us this reputation was William Dawson, prin-
cipal of McGill from 1855 to 1897.
Dawson was a Nova Scotian who studied geology at the University of
Edinburgh where he put up with the dull lectures much more willingly
than Darwin. On return to Canada he began to publish geological articles
and in 1855, his first book, Acadian Geology, was published. The McGill
University Dawson took over that year was very different from the one
where I work. It had 16 professors, but 15 of them were only on staff part-
time. Dawson played a large part in making the university financially
secure and building its early reputation. Sir William Dawson obviously did
not find the job of establishing McGill very challenging because while
doing this, he also delivered approximately 20 lectures a week on a variety
of subjects, published a series of geological papers and books, and as a
sideline, presumably to fill in his spare time, he tried to demolish Darwin’s
theory of evolution. And that explains how William Dawson wormed
his way into this section of my book, which is not about Darwin, but
about a cacophony of critics. I intend to examine the evolution of anti-
evolutionists.
The first school: early emotionals. As I see it, there are four schools of
anti-Darwinists. Dawson was a holotype of the first of these: the early emo-
204 DARWIN AND EVOLUTIONARY SCIENCE

tionals. This group of Darwin’s critics are easily recognized by their similar-
ity to those Anglo-Saxon tourists who believe that everyone in the world can
understand English provided it is spoken in a sufficiently loud voice. The
early emotionals shouted their opposition rather than learning the language of
their opponents.
The early emotionals were characterized by a clear idea of the way in
which human beings should behave. They saw in Darwinism a dreadful new
religion that would make us deviate even farther from the ideal behaviour.
What Dawson opposed was the idea of a process that was not entirely and
continually guided by a divine intelligence.
If the universe is causeless and a product of fortuitous variation and selec-
tion, and if there is no design or final cause apparent in it, it becomes liter-
ally the enthronement of unreason, and can have no claims to the venera-
tion or regard of an intelligent being. If man is merely an accidentally
improved descendant of apes, his intuitions and decisions as to things
unseen must be valueless and unfounded. Hence it is a lamentable fact that
the greater part of evolutionist men of science openly discard all religious
belief, and teach this unbelief to the multitude who cannot understand the
process by which it is arrived at, but who readily appreciate the immoral
results to which it leads in the struggle for existence or the stretching after
material advantages. It is true that there may be a theistic form of evolu-
tion, but let it be observed that this is essentially distinct from Darwinism
or Neo-Lamarckianism. It postulates a Creator, and regards the develop-
ment of the universe as the development of His plans by secondary causes
of His own institution. It necessarily admits design and final cause.
(Dawson 1890)
Dawson tries to convert us, largely by emotional attacks like this, to show
that the theory of evolution will have a short life and is already dying. Lest I
be unfair to Dawson, let me acknowledge that he also had some scientific
objections. He recognized, for example, that no case of transformation of
species had been observed, that the theory did not account for the origin of
sex, and that a mechanism was required to produce variance, but the only
one postulated, inheritance of acquired characteristics, had been falsified by
August Weisman. However the stronger element in Dawson’s anti-Darwinism
is the appeal to emotion.
Dawson was not a solitary representative of this school. He had some
unusual allies, one of the most surprising being the dogmatic atheist George
Bernard Shaw. Shaw hated evolution. He, like Dawson, had a vision of what
the human race should be, and he saw evolutionary theory as something that
opposed his lifelong battle to cure all of the social and intellectual sickness of
mankind.
205CRITICS OF DARWIN

Shaw attacked Darwin on a number of occasions, the most vicious being
in the preface to Man and Superman (1931).
I really do not wish to be abusive; but when I think of these poor dullards,
with their precarious hold of just that corner of evolution that a black-
beetle can understand — with their retinue of twopenny-halfpenny
Torquemadas wallowing in the infamies of the vivisector’s laboratory, and
solemnly offering us as epoch-making discoveries their demonstrations
that dogs get weaker and die if you give them no food; that intense pain
makes mice sweat; and that if you cut off a dog’s leg the three-legged dog
will have a four-legged puppy, I ask myself what spell has fallen on intel-
ligent and humane men that they allow themselves to be imposed on by
this rabble of dolts, blackguards, imposters, quacks, liars, and, worst of all,
credulous conscientious fools. Better a thousand times Moses and Spur-
geon [a then famous preacher] back again. After all, you cannot under-
stand Moses without imagination nor Spurgeon without metaphysics; but
you can be a thorough-going Neo-Darwinian without imagination, meta-
physics, poetry, conscience, or decency. For “Natural Selection” has no
moral significance: it deals with that part of evolution which has no pur-
pose, no intelligence, and might more appropriately be called accidental
selection, or better still, Unnatural Selection, since nothing is more unnat-
ural than an accident. If it could be proved that the whole universe had
been produced by such Selection, only fools and rascals could bear to live.
In the preface to Back to Methuselah, Shaw (1921) provided his longest
attempt to discredit evolutionary theory. Here we see why Shaw hated it so
much.
There is a hideous fatalism about it, a ghastly and damnable reduction of
beauty and intelligence, of strength and purpose, of honour and aspiration.
Shaw believed that the theory of evolution reduced human virtues to chance
events. And what is wrong with chance events? Shaw says that if they rule us:
What hope is there then of human improvement? According to the Neo-
Darwinists [and] to the mechanists no hope whatever, because improve-
ment can come only through some senseless accident, which must ... be
presently wiped out by some other equally senseless accident.
Shaw also rustled up the odd scientific argument, but his main thrust was
emotional. He ridiculed evolutionists and tried to defeat them by demonstrat-
ing his intellectual superiority.
Biologists withstood the attack. The early emotional school is now virtu-
ally dead. As expounded by fundamentalist Christians, it still serves a role in
206 DARWIN AND EVOLUTIONARY SCIENCE

allowing novices in evolutionary theory a chance to fight against a weak foe,
and so sharpen their intellectual teeth. If this opposition is often a match for
us, it is a measure of the over-confidence scientists put in their positions and
a warning about the need for thought and preparation of even a clear-cut
case.
The second school: directional deists. The directional deists constitute a
more formidable and organized opposition to Darwin than the early emotion-
als. This is because they not only bring a wider range of scientific objections
to bear on Darwinian theory, but also because they are more constructive.
They do not just destroy Darwinian theory. They provide an alternative.
This is essentially a French school of thought, derived from Lamarck’s
early theory. Lamarck postulated that when God created the earliest single-
celled creatures, He built into them a directive force that influenced the direc-
tion of evolution. Thus evolution according to Lamarck is not a random,
directionless process. There may be random elements in it caused by unusual
interactions of a species with its environment, but the mainstream of evolu-
tion inexorably flowed towards a god-like creature: Homo sapiens. The three
best-known directional deists are, in order of appearance, Henri Bergson,
Pierre Lecomte de Noüy and Pierre Teilhard de Chardin.
The first of the three is particularly interesting. Bergson (1911) disliked
evolution, not for its own sake, but because he thought it supported determin-
ism. He wanted to believe that human beings have free choice. He disliked
Darwinian evolution because he disliked deterministic philosophy and deter-
ministic science. Bergson developed a whole battery of probabilistic argu-
ments to show that the selection of random mutations was simply not power-
ful enough to cause the evolution of highly complex structures. He postulated
the existence of an élan vitale that would drive evolution towards improve-
ment.
The eye particularly fascinated Bergson. He argued that so complex and
perfect a structure as the eye could not evolve by accumulation of chance
mutations, because any mutation would make it less effective. Take this in
conjunction with the fact that the eye and brain must evolve simultaneously
and we are straining credulity. Finally, when we consider that similar eyes of
great perfection evolved independently in vertebrates and in the scallop
(Pecten), we realize the eye could not possibly have arisen by Neo-Darwinian
evolution.
Bergson’s criticism implies that the eye must be perfectly and fully formed
if it is to function. But this is contrary to observation. Comparative anatomy
provides us with a series of photo-receptors ranging from simple light sensi-
tive cells to the complexities of a falcon’s eye, all of which are functional and
useful to their possessors. Similarly, we ought not be surprised that evolution
207CRITICS OF DARWIN

in different lines should converge to one of a few solutions to the same phys-
ical problems: light sensitivity.
Bergson produced a series of impressive arguments of this type but they all
had the same weakness. They were founded on totally non-quantitative statis-
tics. Bergson was a master of the “logically-black-is-white-slide’ (Flew
1975, Peters 1991a) whereby improbable becomes highly improbable and
ultimately impossible.
Lecomte du Noüy (1947), a biochemist, made his contribution by over-
coming this weakness of qualitative argument. He worked on the origin of life
and demonstrated — to his own satisfaction — that for a single protein
molecule to have originated by chance, within the lifespan of our universe,
we would have needed a universe entirely made of amino acids, 10
19
light
years in diameter. He also developed an argument from the second law of
thermodynamics to the effect that because the universe tends to disorder, evo-
lution to greater order is impossible.
Both of du Noüy’s objections now seem easy to dismiss. Arguments about
the probability of an observed event depend on the null model one has in
mind; whether an event seems probable or not depends on the selected model.
We need only posit rules for the assembly and duration of amino acids to
make the events that are improbable to du Noüy probable to an evolutionist,
and there is now experimental evidence that the latter position is more cor-
rect. Similarly, no one is any longer puzzled by local increases in complexity
in parts of an open system like the biosphere: the solar system is tending to
greater entropy and the biosphere simply slows this flux temporarily. Biolog-
ical complexity is an eddy in the heat death of the universe.
Finally, Teilhard de Chardin (1955, 1959) does not attack the old, but
creates anew. For Bergson’s élan vitale, he substitutes a universal mind-
substance that has God-given tendency to coalesce. Evolution is proceeding
to “point Omega” when all the mind in the universe will coalesce in perfect
unity with the mind of God. Needless to say, we have not reached point
Omega yet, but it does provide a goal for the universe, and that is apparently
a bodily need for this school of critics. They are vitalists who introduce an
unobservable into science and thereby make the universe more understand-
able, but no more predictable.
The third school: cataclysmic creationists. The catastrophists have a
long honourable lineage that can be traced back to Buffon and Cuvier. Their
argument is that there has not been enough time for Darwinian evolution to
produce the vast array of highly perfected species we see on earth. To speed
up the process, they postulate a series of cataclysmic events that eliminated
most existing species, followed by either a new supernatural act of creation or
a sudden evolutionary spurt.
208 DARWIN AND EVOLUTIONARY SCIENCE

The most esteemed of the cataclysmic creationists was Louis Agassiz.
Agassiz, like Dawson, was a geologist and a powerful academic. He is best
known as an advocate of the theory of the ice age and as a teacher at Harvard
University.
Agassiz argued until he died that the facts falsified evolutionary theory
and fitted special creation. He believed that God had created four distinct
groups of animals at the beginning: chordates, molluscs, articulates and ver-
tebrates. Agassiz (1859) held that these four groups have no anatomical sim-
ilarities and thus offer no evidence of evolutionary connections. And as God
creates new species, he always follows one of these four basic plans. Since
the four coexist everywhere, the effect of environment on animals must be
negligible.
A more recent cataclysmic creationist is Immanuel Velikovsky. He is dis-
tinct from all other cataclysmic creationists in that God is missing from his
scheme. Velikovsky’s cataclysms were produced by natural events. Briefly,
he believed that planetary near-collisions so dislocated the earth that the con-
ditions of life on earth changed dramatically and instantaneously (Chap-
ter III). These changes caused mass extinctions, but instead of postulating
supernatural acts of creation, Velikovsky postulated that the great heat and
intense radiation associated with his cataclysms would cause multiple muta-
tions and instantaneous formation of new species. Unfortunately, Velikov-
sky’s genetic ideas are presented so summarily that it is not clear how these
new species arise.
Although they differ greatly in matters of detail, the catastrophists agree
on one point: they reject uniformitarianism, and hold that we cannot explain
the facts of life unless we postulate the existence of unobserved phenomena
such as world-wide cataclysmic events. This school has perhaps gained new
respectability in the last decade or so, because paleontologists are increas-
ingly willing to entertain the idea of catastrophes and variable evolutionary
rates. However, contemporary ideas about catastrophes require no fundamen-
tal change on the part of the scientific community and have been incorporated
into evolutionary thought relatively easily. They differ from their predeces-
sors in using the full scope of geological time and so need no recourse to spe-
cial creation or novel causes of evolutionary change.
The fourth school: Popperian purists. The last group of critics of evolu-
tionary theory are a strangely schizophrenic lot because they do not reject
evolutionary theory. Neither do they try to replace it with another
theory. Deep in their hearts they believe in evolutionary theory as if it were a
truth.
Let me describe these Popperian purists to you. Their name comes from
their prophet: Sir Karl Popper, a philosopher of science. He is largely respon-
209CRITICS OF DARWIN

sible for convincing us that we cannot verify a theory; we can only falsify it.
Thus Popper holds that science progresses by discovering mistakes and cor-
recting them, not by establishing truths. Scientists do this by testing the pre-
dictions of their theories. It is a corollary of this view that theories are rated
not by their truth value, but by the richness of their predictions. A good theory
makes many predictions about different categories of facts and consequently
offers us many opportunities to falsify it. A poor theory makes predictions
about a small body of facts. If a statement makes no predictions, it is not a
theory at all.
To look at Popper’s arguments another way, what a scientific theory really
tells us is that, given present circumstances and knowledge, one or more
specifiable states will not be observed. The best theory rules out all but one
future observation. It is easily falsified. The worst theory rules out no future
state; so whatever happens is consistent with the theory. Nothing can falsify
such a bad theory.
I can illustrate these differences with a simple example. The theory that
“on April 1, the sun rose at 7:06 AM south-east of Montreal” is a good
theory. The theory that “the sun rose on April 1” is less good. And the
theory that “either the sun rose sometime in an unspecified direction from
somewhere, or it did not” is such a bad theory that it is no theory at all. The
Popperian purist judges a theory only by its value as a predictive tool. I have
purposely set this example in the past to stress that the theory predicts an
unknown state, not simply a future state. And I have chosen this familiar
example to stress that the knowledge we gain is personal knowledge, not
just the realization that someone else knows the answer. Alternatively one
may say that the future which a theory predicts is the future state of our
knowledge.
Not all Popperian purists are anti-evolutionists. Those who are believe
that the theory of natural selection is no theory at all because it makes no
predictions that we can falsify. Most are not so extreme. They are like the
agnostic who is desperate to believe, but argues against the existence of God
in the hope that someone will provide the conclusive evidence that She
exists. They argue against the theory of evolution by natural selection, but
hope that their argument will one day attract an evolutionist who can con-
clusively demonstrate that the theory makes falsifiable predictions. This is
my position.
I claim to be a Popperian purist, but I am very close to agnosticism. My
childhood conditioning and undergraduate training were so effective that in
the innermost, animal reaches of my brain, where there is no science, there is
the knowledge that the theory of evolution by natural selection is true. Unfor-
tunately, I cannot show this to be so.
210 DARWIN AND EVOLUTIONARY SCIENCE

Two other biological schools. There are other kinds of biologists. Bio-
logical historicists see science as a narrative describing the sequence of
causal events that preceded any observation, and the true chronicle of events
that preceded the present on earth. They are fascinated by evolution, but are
more interested in explaining past observations than predicting new ones. If
they acknowledge the importance of prediction at all, it is to justify the expla-
nation they have already developed.
A sixth group of biologists I will call the heurists. They see science as a
way to inspire themselves to create more theories or to realize a unity with
nature that they call understanding. For example, many Darwinians and Neo-
Darwinians try to re-trace the thoughts of their name-sake. They hope that
this vicarious recreation of Darwin’s mind will inspire them to similar under-
standing and creativity. If heurists accept that prediction is important, it is
because they see prediction as a useful way of warranting their understand-
ing. If their understanding is sound, it should lead to predictive success, but
whether that prediction helps us gain control over our lives is incidental.
Conclusions
What is the message of this re-examination of old arguments and observa-
tions? In part, it serves to remind the reader of a tradition of anti-Darwinism
that dates back to the last century, and to suggest that not all of this resistance
is rooted in fundamentalist dogma. There are alternative ways of viewing
science. A university program in biology should offer a wide assortment of
historicists, heurists, and Popperian Purists. The student would then have the
opportunity to sample all points of view, and come out of the system behav-
ing more like a scientist than like a Pavlovian dog. I hope these students,
better trained and more aware than I was, will think more about the nature of
science than I did, and so avoid the slavish conditioning that still restricts my
thought.
211CONCLUSIONS


XV Is The Future Grim?
“A man’s reach should exceed his grasp, else
what’s a heaven for?”
Robert Browning
[Andrea del Sarto (1855)]
For most university biologists, science remains a glorious entertainment. For
them, calls to change or to rethink the foundations of research seem unneces-
sary. After all, they can pursue their profession in the same way as their pre-
decessors, earn a decent wage, enjoy the respect of their peers, and sometimes
make a contribution towards the sum of human happiness. Most of us want
little more.
This chapter suggests that, pleasant as a life in science may have been,
those who imagine it will not change are living in a fool’s paradise. I believe
that society feels the time has come to change the university. If university
scientists were to think about science, we too would see that a change is in
store. If professors do not lead the way in this process, society may force us
to accept changes that are both more onerous and less productive.
Any imposed change threatens our ideals as professors. These are to mas-
ter the current ideas of humanity, to contribute our own ideas to this precious
store, and to disseminate both. There is little else that justifies our place in
society. Indeed, because ideas are the most lasting of human accomplish-
ments, there may be nothing else that matters in life, but ideas. University
staff are therefore in a rarely privileged position. We are the pampered
guardians of the intellectual traditions of humanity.
The best ideas in science are theories. Thus our efforts as university scien-
tists are directed to the assessment of existing theories and the production of
new ones. As idealists, we should encourage changes that favour theory, but
inflexibly oppose any change that makes theorizing more difficult. Any
changes to the university should be intended to produce, in our lives and in
the lives of our graduate students, the conditions that are most conducive to
theory-making and theory-testing.
The Gilt Age of University Research
After World War II, an awe-struck society held an unrealistically exagger-
ated view of the collective prowess of its scientists. Science, particularly the

pure science of the universities, was touted as a panacea. University scientists
were consulted on all important matters, quoted in the press, invited to advise
governments, and raised up in the social hierarchy. During this period, we dis-
covered a social conscience. Staff lectured to schools, associations and politi-
cians. We joined extra-mural committees and spent time consulting with
industry or working on government commissions.
The postwar enthusiasm for universities convinced us of our greatness. It
convinced us that we could be all things to all people and that in us alone lay
the power and knowledge to save the world. We would solve practical prob-
lems, teach everything to the new generations and preach to the community at
large. We would discover truth, and our success would demonstrate the supe-
riority of intellect over materialism. These beliefs made every two-bit depart-
ment aspire to greatness.
Not only were we wooed rapidly into accepting society’s inflated opinion
of universities, but we also convinced ourselves that these remarkable powers
would, if provided with the proper nourishment and exercise, continue to
grow. Young turks dismissed the aging scholars in cluttered labs as irrelevant.
They talked not of a science they loved, but of developing excellence, import-
ing eminence, capturing the very best of the young Ph.D.’s, and building
inter-disciplinary teams. We ridiculed the short-sighted granting agencies that
did no more than keep us salivating with slow infusions of thin gruel, and
were encouraged to seek new, more generous sources. We pressured the
library for our share of new journals and symposia volumes, and the univer-
sity for new buildings and research facilities. At one time, we even put six
graduate students in the space designed for three.
Meanwhile, growth of the university and the egocentricity of its staff
caused committees to proliferate and administrative duties to become more
onerous. Within my department alone over twenty committees flourished. I
could go on to recount our forays into big science and our discovery of multi-
disciplinary institutes, but these stories are familiar. We had the money and
time to do more and more, and we did. All these extra duties left little time for
reading. To keep up with science we became world travellers, attending sci-
entific gatherings at up to three exotic places a year.
The Gathering Challenge to University Science
Adulation, like power, corrupts. Our particular form of corruption was
that we came to believe what the newspapers said about us. We could do
anything provided we had enough money. The money came (we could and
did say even that was not enough), the years passed, but we did not pro-
214 IS THE FUTURE GRIM?

duce the promised successes. Our disillusioned bride realized our failure
first and now we too suspect the truth. We had not fully recognized our
limitations.
The postwar honeymoon of the university and the public was primarily
due to the widespread belief that a university degree was the surest key to
the good life for society and personal success for the individual. Student ac-
tivism, devaluation of the degree designation by mass education, and then
economic realities destroyed that faith; and a more realistic evaluation of the
material value of university degrees has decreased teaching allocations. A
second problem for university science is a growing suspicion that much of
the work scientists define as important is actually irrelevant and that useful
scientific output per dollar may be unrelated to the dollars per scientist.
Governments have already reacted to these changes by reducing support for
universities.
I doubt that our modern troubles arose simply because society sometimes
wavers in its support for science. Sometimes, society adopts new patterns. I
think the latter is happening now. Society has permanently changed its atti-
tude to universities, and that changing temper of society will inevitably
change the conditions of university science. The new trend is not a local
Canadian problem. It permeates the United States, Britain, Germany and
other nations because it reflects a fundamental limitation of resources. As
new patterns form in society, some old institutions suffer. I fear the university
is due for a little suffering.
We should expect our budgets to become still less generous. Science has
grown exponentially since the 17th century. Its doubling time is about 10 to
15 years, twice as fast as the economy and three or four times faster than the
population of North America and Europe (Chapter I; Price 1986). This can
only be a temporary situation. Eventually, the growth rate of science must
decrease until it comes into line with the economy and the population.
Although we cannot predict the final share of society’s resources and person-
nel that science can expect, this decrease must make itself felt sometime
within the next 100 years. By then, extrapolation of historical rates of growth
show that science would be as large as the economy and that the population
of scientists would exceed the number of people (Price 1986). This Malthu-
sian argument against the continuing unrestricted growth of science, com-
pounded by the disillusionment of society with university science, leads me
to suppose that the financial pinch in which we now find ourselves is no tem-
porary correction. It is a permanent change.
Since we can expect further belt-tightening, ‘Science Policy’ will continue
to evolve in its present direction. We ought to expect more economic
restraint, not a return to opulence. Tighter control over research monies
215THE GATHERING CHALLENGE TO UNIVERSITY SCIENCE

means these monies will be redirected from universities to government agen-
cies where costs and research can be better controlled. Even more emphasis
will be placed on contract work, industrial and government partnerships, and
on group research into subjects of direct interest to government agencies.
There will be less call for independent theory-making, and pure research by
university staff will be further downgraded. Graduate students, who now per-
form the bulk of university research, will be less in demand and many of
those who do enter graduate school will work on applied research, not
science. If this scenario is even approximately correct, the attitudes we have
fostered over the last 50 years may have to be abolished.
University Responses
We professors must respond to government and societal penny-pinching.
But how? Should we spend more time in public relations, shouting our
praises? Should we lobby for a larger share of the tax revenues? Should we
try to convince the appropriate agencies that because our particular university
is better than others, it needs a bigger share of the budget for post-secondary
education?
Before we spring to the accustomed defensive position of every privileged
minority, we should ask whether or not this reduction in status and financial
support really will decrease our ability to do good science. I think it could, but
only if we respond inappropriately. I also think it has given us the opportunity
to re-evaluate ourselves and to do better science.
I had six years experience with big science while directing the Char
Lake Project which was but a trivial part of the massive International
Biological Program. This experience convinced me that big money makes
administrators of scientists and produces data but not ideas. I believe that
big science restricts, rather than expands, our mental horizons. In our uni-
versity role, we should exercise our talents to produce ideas, not harassed
executives.
An inappropriate response to shrinking budgets will simply dissipate our
energies striving for funds to maintain our research budgets and defending
our own area in frantic turf wars with other researchers. If I thought that spir-
ited defense of our present position would succeed and that success would
help us achieve our ideals, I would counsel battle. But we would not succeed.
Battles must sometimes be fought, but one should always avoid a battle when
the outcome will be disastrous. There is no gain for us in a protracted defense
of the big science we have come to practice.
216 IS THE FUTURE GRIM?

Even if we fought and won a momentary respite, I am uncertain that a
return to fat budgets and more-of-the-same would achieve our ideals. The
universities could perhaps find a new functional role for themselves that
would justify a bigger share of the pie, but our concern is to protect our old
role, the preservation and development of ideas, not to acquire new duties.
And in any case, the Malthusian restriction, that we cannot indefinitely grow
faster than our resources, would make itself felt again in short order.
My colleagues and I already spend our days and nights racing to keep up
to our image of what a scientist should be. We cannot fight harder, indeed
most of us have already compromised ourselves by trying to do too much.
Deep down, we know we are frauds (Medawar 1990). University scientists
are simply normal men and women doing our best to keep our heads above
the flood of duties, demands and interests that only a superman could do well,
but afraid to admit that we can only do them badly. Perhaps we will offer a
secret sigh of relief if society takes away our Captain Marvel cloaks and
forces us to consider where we go from here.
The cure I recommend involves no genius, no technological break-
throughs, nor any other mythical unicorn horn. It is simply the ecological
solution that we must learn to live with less: less waste, less heat, less ineffi-
ciency and less ineffectiveness, but not with fewer ideals or fewer ideas. We
should seek out any change that will lower our aspirations until they come
within shouting range of our abilities. I simply recommend that we return to
our ideals.
A Policy for the Future: Closing the Aspiration Gap
There is now so large a gap between what we can do and what we are
expected to do that a sense of frustration and impotency is inevitable. A gap
may be essential, but a chasm is counter-productive. I suggest that we deter-
mine what it is we must do and try to find the most efficient way to achieve
those ends. To do so, we must sacrifice many of the trappings we have accu-
mulated in the last generation, but we need not compromise our effectiveness
in learning, teaching and developing theory. All of my recommendations are
therefore intended to produce more uninterrupted time for thought and
research without spoiling our teaching efforts.
Undergraduate teaching. The equilibrium between teaching and re-
search is unstable, so there can be no optimum balance between these two
critical activities of a university professor. Nevertheless, we must recognize
that we are paid to teach, and that everything else is largely self-imposed. In
217A POLICY FOR THE FUTURE: CLOSING THE ASPIRATION GAP

keeping with our loss of prestige, we can expect society to stress the impor-
tance of undergraduate teaching. Since there will be no decrease in student
numbers, no increase in staff, and continued demands for good teaching, we
must make our teaching much more efficient and effective.
Equalize teaching loads. Assuming that a happy teacher is a better teacher,
we should equalize teaching loads. This will make teachers happy because
the greatest source of friction in most departments is the perceived inequality
of the commitments of various colleagues to the burden of undergraduate
instruction. The reassignment of teaching loads must be done without refer-
ence to outside duties, to the number of pages of print we produce annually,
or to the number of graduate students we train. The workload should be
evenly distributed. This will protect those who enter the system in the future
from being pushed into too much teaching, but it will also help to close the
aspiration gap by impressing us with the fact that we are here as teachers and
we should not be rewarded for other activities by having our teaching load
reduced.
Abolish tenure. It is a corollary of the central role of teaching in university
life that we should be judged as teachers and that useless teachers should be
dismissed. This freedom can only be achieved if we abolish tenured posi-
tions. I see no virtue in cut-throat competition, so some alternatives to throw-
ing our colleagues into the street must be found. Perhaps those who fail as
teachers too late in life to find another job could be transferred to administra-
tive posts.
Tenure is widely applauded as a bastion protecting those whose views
challenge authority. Thus, the abolition of tenure is abhorred because it would
leave the most precious members of the community open to arbitrary dis-
missal. The protection of deadwood is seen as the cost of academic freedom.
This defense seems disingenuous: professors with unpopular views have
never been well defended by tenure, because university and state administra-
tions have too often buckled to public pressure. Tenure really protects incom-
petence.
The end of tenure will impress on us the fact that we are ordinary mortals,
not above judgement. It will also make it easier for new staff to become good
teachers, because they will justifiably devote more time to preparation for
teaching. Neither will they have to do research in desperate haste to achieve
the number of publications demanded by tenure committees. They can do it
because they enjoy it.
Reduce course offerings. The equalization of teaching loads could stifle
research, unless we are careful to contain the total teaching burden. Therefore
I recommend that we reduce the number of course offerings by pooling
specialized courses and allowing students more freedom of choice among
218 IS THE FUTURE GRIM?

courses and subject matter. This reduction would be facilitated by eliminating
labs from many courses, and limiting the number of lecture hours, thus giving
students more time for independent work.
We should also eliminate all courses with less than 10 to 30 students. The
cut-off is arbitrary but a minimum class-size acknowledges that small courses
waste money and professorial time. If offered at all, they should be offered
only every other or every third year. Small, specialized courses are in demand
by staff who see in them the opportunity to study their own speciality in even
more detail and to teach an easy subject. They are defended as a device to
attract graduate students, but obviously this is not so. If it were, enrolments
would be larger.
Reduce teaching burdens. To make our work as efficient as possible, we
should eliminate or at least limit the mind-numbing chores that seem so much
a part of university teaching. We should exploit the multi-media approach, the
computer and any other approach or device that will eliminate repetition of
lectures and hand-marking of anything over 100 identical examinations in a
week or two by one professor. Our objective should be to make teaching large
classes an enjoyable job.
Graduate training. Graduate education is also one of our primary duties.
I have de-emphasized it in my considerations because graduate education will
be less important in the university science of the future. Decreased budgets,
increased emphasis on undergraduate education and the de-emphasis of inde-
pendent research will relieve us of the obligation we once felt to increase our
graduate training programme. Indeed, a slower rate of growth of science will
mean that our graduate training programmes will have to be curtailed. Pro-
fessors will therefore have more time and will be able to do the work them-
selves.
To encourage this curtailment, we should institute a policy that will pro-
tect us from our own inability to refuse good applicants. Even good graduates
cost money and thus distract professors from research by sending them out
looking for more or bigger research grants. Weak students do this, and also
demand even more professorial time in training.
Scholarship. Scholarship is what we modestly call “our own work”. My
schemes are intended to make it possible for us to do and enjoy our own work
in the coming university environment. We must find ways to avoid the busy
work that cuts us off from research.
For example, the telephone is probably the single greatest obstruction to
scholarship and meaningful intercourse amongst staff. I would remove all
telephones from professors’ offices, although I would not prohibit graduate
students or staff from renting their own. Fax machines and E-mail are simi-
larly invasive.
219A POLICY FOR THE FUTURE: CLOSING THE ASPIRATION GAP

The mobility of big science also robs us of the calm we need to think and
to develop theory by bringing us colleagues and visiting lecturers who them-
selves dedicate days of their time to deliver an hour’s lecture. Too often, that
lecture is appreciated by only a tiny fraction of the audience. The rest are
there only to hold up the side, or as a courtesy to the visitor. This is clearly
inefficient. All visiting lecturers and seminars should be aimed at undergrad-
uates first and graduates second, since students more frequently are the
people with the time to listen to and the minds to learn from visitors.
Inter-disciplinary institutes are also devices to sabotage meaningful
research. They are based on a flawed model of our needs and abilities
(Chapter XIII) and they lock the individual into the narrow confines of his or
her speciality as one element in a systems approach (Chapter VIII). If one
wishes to develop one’s own ideas in science, one must not start such insti-
tutes or participate in their programmes.
Staff members should be actively discouraged from seeking large re-
search grants. They might, for example, be asked to justify their proposals
in terms of their ideals to the department staff. Large research grants in-
crease the output of data, but spending money is such a time-consuming
occupation that it can reduce the quality of research. The danger will in-
crease in the future because the evolving science policy will mean that when
we get large grants, we will be required to conduct research programs de-
signed more by others than by ourselves. All of this is obviously anathema
to our ideals as professors.
Administration. Administration presents a problem for which I have no
solution. At the departmental level, we have some control over the amount of
administrative busy-work we impose on ourselves, but we have none over the
university as a whole. Thus, university committees and commissions will
continue to propagate regardless of our attitude toward them. Furthermore,
we will be expected to serve on them.
My limited experience with these activities is that they waste a great
deal of time. They are unnecessarily protracted by aspiring administrators
who cannot bear the thought of disappearing back into the obscurity of
their own office, whereas any professor worthy of the name should be eager
to return to the lab or office to pursue his or her ideas. Although we cannot
escape the feeling of obligation to contribute our minimal share towards the
running of the university, it would probably function just as well if we were
to opt out.
Perhaps there is nothing we can do about this impediment to teaching and
scholarship, but we should disabuse ourselves of the belief that active partic-
ipation in university affairs is a virtue. Certainly, administrative duties should
be irrelevant to advancement and job security at the university.
220 IS THE FUTURE GRIM?

Two reservations. What I have tried to say is that for almost half a cen-
tury, we have been obsessed with a chimera: we have tried to achieve better
science with grantsmanship instead of scholarship. I have made entirely rea-
sonable suggestions like these in the past and been surprised that they were
not universally accepted. My listeners instead suggested that such changes
could undermine science by setting such low goals that researchers would
cease to strive for new ideas and become apathetic towards science. We
would return to a tweedy and unproductive past. From these objections, I can
only infer that my cloudy literary style can obscure the dazzling brilliance of
my ideas. Therefore, I will end with a brief rebuttal of those suggestions.
A formula for apathy? There is a widespread belief that because we have
an animal need to strive, it would be undesirable for us to reach our ideals.
This fear is now particularly acute, because apathy is the disease of the age. It
is not my position that we should stop striving. That would merely aggravate
the situation and produce more apathy. The modesty of my proposals could
inadvertently discourage some scientists and prevent them from trying to
improve, but that is not my intent.
Unlike Browning’s Andrea del Sarto, I believe it is more frustrating to
attempt the impossible than to set our goals so low that we can actually reach
some of them. If we actually did achieve our modest goals, we would not be
finished as scientists. Success would stimulate, not inhibit us.
The goals of a scientist are like those of the exhausted traveller. It is not
the end of the journey, but the next tree or stone that remains the goal. This is
the view of science propagated by 20th century philosophers like Sir Karl
Popper and historians of science like T. S. Kuhn (Chapter III). We never reach
an ultimate truth. We merely continue to elaborate and change our ideas as we
progress, not to a lasting goal, but farther and farther away from our primi-
tive, fumbling beginnings. Thus I think we can be both realistic and idealis-
tic. By combining the two, we will stimulate not inhibit scientific excitement.
A step back? For some, my suggestions represent a step back to some lower
rung of the evolutionary ladder of science, but that applies only for some
unessential aspects of research. I am looking back or down only if it is retro-
gressive to emphasize doing science rather than chasing research dollars. For
a generation, university scientists aspired to greatness, and almost destroyed
themselves. Individuals competed, unified departments fragmented, and some
of us discovered that university life was no longer pleasant. In this sense, I
hope my solutions are retrogressive.
I want to escape what the modern university has become. However, I am
not prepared to return to mindless, unchallenging research that grubs for new
facts or defends favourite theories from scientific scrutiny. The opportunity
that social change offers us is not a return to the trifling puzzles of the past,
221A POLICY FOR THE FUTURE: CLOSING THE ASPIRATION GAP

but the chance to consider what we have done, what we are doing, and what
we should do. If we can escape from the trap of our own self-image, we can
begin to contemplate science.
Conclusion
Big science and societal power have caused professors to lose track of
their ideals and goals. In doing so, they have almost destroyed the university
as an intellectual retreat. When I begin to hear more staff members whistling
in the corridors or humming in their labs, I will be content that we have come
into a happier equilibrium with reality. Under these conditions we will not
only be able to do more effective research, but we will also discharge our duty
to society. We will pass on the tradition that intellectual activity is more excit-
ing than material consumption. We will have learned to contemplate science.
222 IS THE FUTURE GRIM?

Acknowledgements
I thank the many individuals who have helped so much in the production of the book.
The Rigler family helped transcribe and edit the lectures. Andréa Grottoli Everett drew
most of the figures; Greg Scarborough and Gary Bowen have been invaluable in the
final stages. I am deeply indebted to David Currie of the University of Ottawa, John
Downing of the University of Montreal, Don Kramer of McGill and to Yves Prairie and
David Bird, both of the Université de Québec at Montréal, for their comments on ear-
lier drafts of the manuscript.
The passage that serves as a prologue is from Molloy by Samuel Beckett, translated
from the French by Samuel Beckett and Patrick Bowles, copyright 1950 and copyright
this translation 1955, 1959, 1966, 1971, and 1976 by Samuel Beckett. Reproduced by
permission of the Beckett Estate and the Calder Educational Trust, London. Rigler
played a reading of this passage by Cyril Cusack at the beginning, and again at the end,
of his courses in history and philosophy of science at both the University of Toronto and
McGill University.


References
Abrams, P. 1983. The theory of limiting similarity. A. Rev. Ecol. Syst. 14: 359–376
Agassiz, L. 1859. An Essay on Classification. Longman, Brown, Green, Longmans &
Roberts, and Trubner, London
Ahrens, M. A. and R. H. Peters. 1991. Plankton community respiration: relationships
with size distribution and lake trophy. Hydrobiologia 224: 77–87
Allen, T. F. and T. B. Starr. 1982. Hierarchy: Perspectives for Ecological Complexity.
University of Chicago Press, Chicago
Ambühl, H. 1960. Die Nährstoffzufuhr zum Hallwilersee. Schweiz. Z. Hydrol. 22:
564–597
Anonymous. 1972. A record of success. Pensée 2: 11–15
APHA Standard Methods for the examination of water and wastewater. 1989. L. S.
Clesceri, A. E. Greenberg and R. R. Trussell (eds.). 17th edition. American Public
Health Association, Washington, D.C.
Armstrong, F. A. J. and D. W. Schindler. 1971. Preliminary chemical characterization
of waters in the Experimental Lakes Area, northwestern Ontario. J. Fish. Res. Bd
Can. 28: 171–187
Atkins, W. R. G. 1923. The phosphate content of fresh and salt waters in its relationship
to the growth of algal plankton. J. mar. Biol. 13: 119–150
Aure, J. and A. Stigebrandt. 1990. Quantitative estimates of the eutrophication effects
of fish farming on fjords. Aquaculture. 90: 135–156
Bacon, F. 1621. The Advancement of Learning: Novum Organum. (1930 edition.)
Oxford University Press, London
Barica, J. 1984. Empirical models for prediction of blooms and collapses, winter oxy-
gen depletion and a freeze-out effect in lakes. Summary and verification. Verh. int.
Verein. Limnol. 22: 309–319
Barzun, J. 1964. Science: The Glorious Entertainment. University of Toronto Press,
Toronto
Beatty, J. 1980. Optimal-design models and the strategy of model building in evolu-
tionary biology. Phil. Sci. 47: 532–561
Beck, M. B. and E. Halfon. 1991. Uncertainty, identifiability and the propagation of
prediction errors: a case study of Lake Ontario. J. Forecast. 10: 135–161
Beckett, S. 1950. Molloy. (1955 translation.) Grove Press, New York
Beeton, A. M. 1969. Changes in the environment and biota of the great lakes.
pp. 150–187. In: Eutrophication: Causes, Consequences, Correctives; Proceedings
of a Symposium. National Academy of Sciences, Washington, D.C.
Benedict, F. G. 1938. Vital Energetics: A Study in Comparative Basal Metabolism.
Carnegie Institute of Washington, Washington, D.C.
Benndorf, J. 1987. Food web manipulation without nutrient control: a useful strategy in
lake restoration? Schweiz. Z. Hydrol. 49: 237–248
Bentzen, E. and W. D. Taylor. 1991. Estimating Michaelis-Menten parameters and lake
water phosphate by the Rigler bioassay: importance of fitting technique, plankton
size, and substrate range. Can. J. Fish. Aquat. Sci. 48: 73–83
Bergson, H. 1911. Creative Evolution. (1944 edition.) Random House, New York

Bernard, C. 1860. The Cahier Rouge of Claude Bernard. 1967 translation, H. H. Hoff,
L. Guillemin and R. Gullemin (trans.). In: Grande, F. and M. B. Visscher, 1967,
Claude Bernard and Experimental Medicine. Schenkman, Cambridge
Bird, D. F. and J. Kalff. 1984. Empirical relationships between bacterial abundance and
chlorophyll concentration in fresh and marine waters. Can. J. Fish. Aquat. Sci. 41:
1015–1023
Blackman, F. F. 1905. Optima and limiting factors. Ann. Bot. 19: 281–295
Bodenheimer, F. S. 1953. A History of Biology. Dawson and Sons, London
Bonner, J. T. 1962. The Ideas of Biology. Harper and Row, New York
Box, E. O. 1981. Macroclimate and Plant Forms: An Introduction to Predictive Model-
ing in Phytogeography. Dr. W. Junk, The Hague
Box, G. E. P. 1976. Science and statistics. J. Am. statist. Ass. 71: 791–799
Bray, J. R. and E. Gorham. 1964. Litter production in forests of the world. Adv. ecol.
Res. 2: 101–157
Brody, S. 1945. Bioenergetics and Growth. Reinhold, Baltimore, Md
Brooks, J. L. and S. I. Dodson. 1965. Predation, body size, and composition of plank-
ton. Science 150: 28–35
Brown, J. H. 1981. Two decades of Homage to Santa Rosalia: toward a general theory
of diversity. Am. Zool. 21: 877–888
Brum, G. D. and L. K. McKane. 1989. Biology: Exploring Life. J. Wiley and Sons, New
Yor k
Burns, C. W. 1968. Direct observations of mechanisms regulating feeding behavior of
Daphnia in lakewater. Int. Revue ges. Hydrobiol. 53: 83–100
Burns, C. W. and F. H. Rigler. 1967. Comparison of filtering rates of Daphnia rosea in
lake water and in suspensions of yeast. Limnol. Oceanogr. 12: 492–502
Calder, W. A., III. 1984. Size, Function and Life History. Harvard University Press,
Cambridge
Canfield, D. E., Jr. and R. W. Bachman. 1981. Prediction of total phosphorus concen-
trations, chlorophyll a and Secchi depths in natural and artificial lakes. Can. J. Fish.
Aquat. Sci. 38: 414–423
Canfield, D. E., Jr., J. V. Shiremen, D. E. Colle, W. T. Haller, C. E. Walkins III and M. J.
Maceina. 1984. Prediction of chlorophyll a concentrations in Florida lakes: impor-
tance of aquatic macrophytes. Can. J. Fish. Aquat. Sci. 41: 497–501
Carpenter, S. R., J. F. Kitchell and J. R. Hodgson. 1985. Cascading trophic interactions
and lake productivity. BioScience 35: 634–639
Cattaneo, A. 1987. Periphyton in lakes of different trophy. Can. J. Fish. Aquat. Sci. 44:
296–303
Caws, P. 1969. The structure of discovery. Science 166: 1375–1380
Chamberlain, W. M. 1968. A Preliminary investigation of the nature and importance of
soluble organic phosphorus in the phosphorus cycle of lakes. Ph.D. Thesis, Univer-
sity of Toronto, Toronto
Chamberlin, T. C. 1890. The method of multiple working hypotheses. Science old
series 15: 92. Reprinted 1965. Science 148: 754–759
Chambers, P. A. and J. Kalff. 1985. Depth distribution and biomass of submerged macro-
phyte communities in relation to Secchi depth. Can. J. Fish. Aquat. Sci. 42: 701–709
Clements, F. E. 1916. Plant succession: analysis of the development of vegetation.
Publs Carnegie Instn 242, Washington, D.C.
Cohen, B. 1985. Revolution in Science. The Belknap Press of Harvard University
Press, London
226 REFERENCES

Cohen, J. E. 1991. Size, age and productivity of scientific and technical research
groups. Scientometrics 20: 395–426
Cole, J. R. and Cole, S. 1972. The Ortega hypothesis. Science 178: 368–375
Confer, J. L. 1969. The Inter-relationships among plankton, attached algae and the
phosphorus cycle in artificial open systems. Ph.D. Thesis, University of Toronto,
Toronto
Confer, J. L. 1972. Interrelations among plankton, attached algae, and the phosphorus
cycle in artificial open systems. Ecol. Monogr. 42: 1–23
Connell, J. H. and W. P. Sousa. 1983. On the evidence needed to judge ecological sta-
bility or persistence. Am. Nat. 121: 789–824
Cornett,R. J. 1989. Predicting changes in hypolimnetic oxygen concentrations with phos-
phorus retention, temperature and morphometry. Limnol. Oceanogr. 34: 1359–1366
Cornett, R. J. and F. H. Rigler. 1979. Hypolimnetic oxygen deficits: their prediction and
interpretation. Science 205: 580–581
Currie, D. J. 1990. Large-scale variability and interactions among phytoplankton, bac-
terioplankton, and phosphorus. Limnol. Oceanogr. 35: 1437–1455
Currie, D. J. 1991. Energy and large-scale patterns of animal and plant species richness.
Am. Nat. 137: 27–49
Currie, D. J. and V. Paquin. 1987. Large-scale biogeographical patterns of species rich-
ness of trees. Nature 329: 326–327
Darwin, C. 1876. The autobiography of Charles Darwin and selected letters. F. Darwin
(ed.). 1892, 1958. Dover Publications, New York
Dawson, W. 1890. Modern Ideas of Evolution as Related to Revelation and Science.
Religious Tract Society, London
de Chardin, T. 1955, 1965. The Phenomenon of Man. Harper and Bros., New York
Deevey, E. S. 1940. Limnological studies in Connecticut: a contribution to regional
limnology. Am. J. Sci. 282: 717–741
de Grazia, R. E. Jurgens and L. C. Stecchini. 1966. The Velikovsky Affair. Scientism
vs. Science. University Books, New Hyde Park, New York
del Giorgio, P. and R. H. Peters. 1993. The balance between phytoplankton production
and plankton respiration in lakes. Can. J. Fish. Aquat. Sci. 50: 282–289
Dillon, P. J. 1973. The prediction of phosphorus and chlorophyll concentrations in lake-
water. Ph.D. Thesis, University of Toronto, Toronto
Dillon, P. J. and W. B. Kirchner. 1975. The effects of geology and land use on the export
of phosphorus from watersheds. Wat. Res. 9: 135–148
Dillon, P. J. and F. H. Rigler. 1974a. The phosphorus-chlorophyll relationship in lakes.
Limnol. Oceanogr. 19: 767–73
Dillon, P. J. and F. H. Rigler. 1974b. A test of a simple nutrient budget model predict-
ing the phosphorus concentration in lake water. J. Fish. Res. Bd Can. 31: 1771–1778
Dillon, P. J. and F. H. Rigler. 1975. A simple method for predicting the capacity of a
lake for development based on lake trophic status. J. Fish. Res. Bd Can. 32:
1519–1531
Dillon, P. J., W. A. Scheider, R. A. Reid and D. S. Jeffries. 1994. Lakeshore Capacity
Study: Part I - Tests of effects of shoreline development on the trophic status. Lake
Reserv. Manage. 8: 121–129
Dodson, S. I. 1970. Complementary feeding niches sustained by size-selective preda-
tion. Limnol. Oceanogr. 15: 131–137
Douglas, A. E. 1919. Climatic Cycles and Tree Growth. Vol I. Publs Carnegie Instn
289, Washington, D.C.
227REFERENCES

Downing, J. A. 1981. In situ foraging responses of three species of littoral cladocerans.
Ecol. Monogr. 51: 85–103
Downing, J. A. 1984. Assessment of secondary production: the first step. pp. 1–12. In:
J. A. Downing and F. H. Rigler (eds.). A manual on methods for the assessment of
secondary productivity in fresh waters. Oxford Blackwell Scientific Publications,
London
Downing, J. A. 1991. Comparing apples with oranges: Methods of interecosystem com-
parison. pp. 24–45. In: J. Cole, G. Lovett and S. Findlay (eds.). Comparative Anal-
yses of Ecosystems. Springer-Verlag, New York
Downing, J. A., C. Plante and S. Lalonde. 1988. Fish production correlated with pri-
mary productivity, not the morphoedaphic index. Can. J. Fish. Aquat. Sci. 47:
1929–1936
Duarte, C. M. and J. Kalff. 1990. Patterns in the submerged macrophyte biomass of
lakes and the importance of the scale of analysis in the interpretation. Can. J. Fish.
Aquat. Sci. 47: 357–363
du Noüy, P. Lecomte. 1947. Human Destiny. Mentor, New York
East, R. 1984. Rainfall, soil nutrient status and biomass of large African savanna mam-
mals. Afr. J. Ecol. 22: 245–270
Economos, A. C. 1979. On structural theories of basal metabolic rate. J. theor. Biol. 80:
445–450
Edmondson, W. T. 1969. Eutrophication in North America. pp. 124–149. In: Eutrophi-
cation: Causes, Consequences, Correctives. Proceedings of a symposium. National
Academy of Sciences, Washington, D.C.
Edmondson, W. T. 1970. Phosphorus, nitrogen, and algae in Lake Washington after
diversion of sewage. Science 169: 690–691
Edmondson, W. T. 1972. The present condition of Lake Washington. Verh. int. Verein.
Limnol. 18: 284–291
Edmondson, W. T. and J. T. Lehman. 1981. The effect of changes in the nutrient income
on the condition of Lake Washington. Limnol. Oceanogr. 26: 1–29
Edmondson, Y. T. 1971. Some components of the Hutchinson legend. Limnol.
Oceanogr. 16: 157–172
Einsele, W. 1941. Die Umsetzung von zugeführtem, anorganischen Phosphat im eutro-
phen See und ihre Rückwirkungen auf seinen Gesamthaushalt. Z. Fisch. Hilfs-
wissenschaften 39: 407–488
Eiseley, L. 1973. The Man Who Saw through Time. Charles Scribner’s Sons, New York
Eldredge, N. and S. J. Gould. 1972. Punctuated equilibria: an alternative to phyletic
gradualism. pp. 82–115. In: T. J. M. Schnopf (ed.). Models in Paleobiology. Free-
man, Cooper and Co., San Francisco
Elser, J. J. and S. R. Carpenter. 1988. Predation-driven dynamics of zooplankton and
phytoplankton communities in a whole-lake experiment. Oecologia 76: 148–154
Elster, H.-J. 1958. Das limnologische Seetypensystem, Rückblick und Ausblick. Verh.
int. Verein. Limnol. 13: 101–120
Elton, C. S. 1927. Animal Ecology. Sedgewick and Jackson, London
Elton, C. S. 1966. The Pattern of Animal Communities. Methuen, London
Evans, R. D. and F. H. Rigler. 1985. Long distance transport of anthropogenic lead as
measured by lake sediments. Wat. Air Soil Pollut. 24(139): 141–151
Forbes, S. A. 1887. The lake as a microcosm. Bull. Sci. Ass. Peoria, Illinois. 1887: 77–87
Forsberg, C. 1987. Evaluation of lake restoration in Sweden. Schweiz. Z. Hydrol. 49:
260–274
228 REFERENCES

Frank, P. 1949. Modern Science and its Philosophy. Harvard University Press, Cam-
bridge
Frank, P. 1957. The Philosophy of Science. Prentice-Hall, Inc., Englewood Cliffs, New
Jersey
Fretwell, S. D. 1975. The impact of Robert MacArthur on ecology. A. Rev. Ecol. Syst.
6: 1–13
Fry, F. E. J. 1947. Effects of the environment of animal activity. Univ. Toronto Stud.
biol. Ser. 55: 5–62
Galton, F. 1875. English Men of Science: Their Nature and Nurture. Appleton, New
Yor k
Garfield, E. 1977a. The 250 most-cited primary authors, 1961–1975. Part I. How the
names were selected. Curr. Cont. 1977(49): 5–15
Garfield, E. 1977b. The 250 most-cited primary authors, 1961–1975. Part II. The cor-
relation between citedness, Nobel prizes, and academy memberships. Curr. Cont.
1977(50): 5–15
Garfield, E. 1977c. The 250 most-cited primary authors, 1961–1975. Part III. Each
author’s most-cited publication. Curr. Cont. 1977(51): 5–20
Garfield, E. 1985. Uses and misuses of citation frequency. Curr. Cont. 1985(43): 3–9
Garfield, E. 1986. The 250 most-cited primary authors in the 1984 SCI, Part 1. Names,
ranks and citation numbers. Curr. Cont. 17 (10): 3–11
Gauld, D. T. 1951. Diurnal variations in the grazing of planktonic copepods. J. mar.
Biol. Ass. U.K. 31: 461–474
Ghiselin, M. 1969. The Triumph of the Darwinian Method. University of Chicago
Press, Chicago
Glaser, B. G. 1964. Comparative failure in science. Science 143: 1012–1014
Glazier, D. S. 1992. Effects of food, genotype, and maternal size and age on offspring
investment in Daphnia magna. Ecology 73: 910–926
Gleick, J. 1987. Chaos: Making a New Science. Viking, New York
Gliwicz, Z. M. and E. Siedlar. 1980. Food size limitation and algae interfering with
food collection in Daphnia. Arch. Hydrobiol. 88: 155–177
Godbout, L. and R. H. Peters. 1988. Potential determinants of stable catch in the brook
trout (Salvelinus fontinalis) sport fishery in Quebec. Can. J. Fish. Aquat. Sci. 45:
1771–1778
Gomolka, R. 1975. An investigation of atmospheric phosphorus as a source of lake
nutrient. M.Sc. Thesis, University of Toronto, Toronto
Grahame, K. 1966. The Wind in the Willows. Gosset and Dunlop, New York
Gray, R. D. 1987. Faith and foraging. pp. 69–140. In: A. C. Kamil, J. R. Krebs and H.
R. Pulliam (eds.). Foraging Behavior. Plenum, New York
Griesbach, S. and R. H. Peters. 1991. The effects of analytical variations on estimates
of phosphorus concentration in surface waters. Lake Reserv. Manage. 7: 97–106
Hairston, N. G., F. E. Smith and L. B. Slodobkin. 1960. Community structure, popula-
tion control, and competition. Am. Nat. 94: 421–425
Håkanson, L. 1991. Ecometric and Dynamic Modelling — Exemplified by caesium in
lakes after Chernobyl. Springer-Verlag, New York
Håkanson, L., T. Anderson and A. Nilsson. 1990. Mercury in fish in Swedish lakes -
linkages to domestic and european sources of emission. Wat. Air Soil Pollut. 50:
171–191
Håkanson, L. and M. Wallin. 1991. An outline of ecometric analysis to establish load
diagrams for nutrients/eutrophication. Envirometrics 2: 49–68
229REFERENCES

Hall, D. J., S. T. Threlkeld, C. W. Burns and P. H. Crowley. 1976. The size-efficiency
hypothesis and the size structure of zooplankton communities. A. Rev. Ecol. Syst.
7: 177–208
Hall, T. S. 1969. History of General Physiology. Vol 2. The University of Chicago
Press, Chicago
Haney, J. F. 1970. Seasonal and spatial changes in the grazing rate of limnetic zoo-
plankton. Ph.D. Thesis, University of Toronto, Toronto
Haney, J. F. 1971. An in situ method for the measurement of zooplankton grazing rates.
Limnol. Oceanogr. 16: 970–977
Haney, J. F. 1973. An in situ examination of the grazing activities of natural zooplank-
ton communities. Arch. Hydrobiol. 72: 87–132
Haney, J. F. and D. J. Hall. 1975. Diel vertical migration and filter-feeding activities of
Daphnia. Arch. Hydrobiol. 75: 413–441
Hanna, M. and R. H. Peters. 1991. Effect of sampling protocol on estimates of phos-
phorus and chlorophyll concentrations in lakes of low to moderate trophic status.
Can. J. Fish. Aquat. Sci. 48: 1979–1986
Hanson, J. M. and W. C. Leggett. 1982. Empirical prediction of fish biomass and yield.
Can. J. Fish. Aquat. Sci. 39: 257–263
Hanson, J. M. and R. H. Peters. 1984. Empirical prediction of zooplankton biomass and
profundal macrobenthos biomass in lakes. Can. J. Fish. Aquat. Sci. 41: 439–445
Hardin, G. 1961. The competitive exclusion principle. Science 131: 1292–1297
Hardy, A. C. 1924. The herring in relation to its animate environment, Part 1. Ministry
of Agriculture and Fisheries. Fishery Investigations Series. 2(7): 1–53
Harmon, L. R. 1961. The high school backgrounds of science doctorates. Science 133:
679–688
Harvey, P. H. and G. M. Mace. 1982. Comparisons between taxa and adaptive
trends: problems in methodology. pp. 343–361. In: King’s College Sociobiology
Group (eds.). Current Problems in Sociobiology. Cambridge University Press, Cam-
bridge
Hemmingsen, A. M. 1960. Energy metabolism as related to body size and respiratory
surfaces, and its evolution. Rep. Steno meml. Hosp. 9: 1–110
Hempel, C. 1962. Explanation in history and science. pp. 9–33. In: R. G. Colodny (ed.).
Frontiers of Science and Philosophy. University of Pittsburgh Press, Pittsburgh
Hermens, J. L. 1986. Quantitative structure-activity relationships in aquatic toxicology.
Pestic. Sci. 17: 287–296
Hickman, C. P., L. S. Roberts and F. M. Hickman. 1984. Integrated Principles of Zool-
ogy. 7th edition. Times Mirror/Mosby College Publishing, St. Louis, Missouri
Holeton, G. F. 1973. Respiration of Arctic char (Salvelinus alpinus) from a high arctic
lake. J. Fish. Res. Bd Can. 30: 717–723
Holling, C. W. 1959. Some characteristics of simple types of predation and parasitism.
Can. Ent. 91: 385–398
Hoyer, M. V. and D. E. Canfield, Jr. 1991. A phosphorus-fish standing crop relationship
for streams? Lake Reserv. Manage. 7: 25–32
Hrbácˇek, J., Dvoráková, M. V. Korˇínek and L. Procházková. 1961. Demonstration of
the effect of the fish stock on the species composition of zooplankton and the inten-
sity of metabolism of the whole plankton association. Verh. Internat. Verein.
Limnol. 14: 192–195
Hull, D. L. 1974. Philosophy of Biological Science. Prentice-Hall, Englewood Cliffs,
New Jersey
230 REFERENCES

Hutchinson, G. E. 1938. On the relationship between oxygen deficit and the productiv-
ity and typology of lakes. Int. Revue ges. Hydrobiol. 36: 336–355
Hutchinson, G. E. 1951. Copepodology for the ornithologist. Ecology 32: 571–577
Hutchinson, G. E. 1957. A Treatise on Limnology. Vol. 1. Wiley and Sons, New York
Hutchinson, G. E. 1959. Il concetto moderno di nicchia ecologica. Memorie Ist. ital.
Idrobiol. 11: 9–22
Hutchinson, G. E. 1961. The paradox of plankton. Am. Nat. 95: 137–146
Hutchinson, G. E. 1966. The prospect before us. pp. 683–690. In: D. G. Frey (ed.). Lim-
nology in North America. University of Wisconsin Press, Madison
Hutchinson, G. E. 1978. An Introduction to Population Ecology. Yale University Press,
New Haven
Huxley, T. H. 1880. The Crayfish: An Introduction to the Study of Zoology. 1977 edi-
tion. MIT Press, New York
Infeld, L. 1978. Why I Left Canada. Reflections on Science and Politics. McGill-
Queen’s University Press, Montreal
Iverson, R. L. 1990. Control of marine fish production. Limnol. Oceanogr. 35: 1593-
1604
Jassby, A. D. and C. R. Goldman. 1974. Loss rates from a lake phytoplankton commu-
nity. Limnol. Oceanogr. 19: 618–627
Jolicouer, P. and A. A. Heusner. 1971. The allometry equation in the analysis of the
standard oxygen consumption and body weight of the white rat. Biometrics 27:
841–855
Juday, C. and E. A. Birge. 1931. A second report on the phosphorus content of Wis-
consin lake waters. Trans. Wisconsin Acad. Sci. 26: 353–382
Jumars, P. A. 1987. Editorial comment: the evolving natural history of a manuscript.
Limnol. Oceanogr. 32: 1011–1014
Jumars, P. A. 1990. W(h)ither limnology? Limnol. Oceanog. 35: 1216–1217
Kalff, J. and H. E. Welch. 1974. Phytoplankton production in Char Lake, a natural polar
lake, and in Meretta Lake, a polluted polar lake, Cornwallis Island, Northwest Ter-
ritories. J. Fish. Res. Bd Can. 31: 621–636
Karl, D. M. and G. Tien. 1992. MAGIC: a sensitive and precise method for measuring
dissolved phosphorus in aquatic environments. Limnol. Oceanogr. 37: 105–116
Keddy, P. A. 1989. Competition. Chapman and Hall, London
Keeton, W. T. and J. L. Gould. 1986. Biological Science. 4th edition. W. W. Norton and
Co., New York
Kibby, H. V. and F. H. Rigler. 1973. Filtering rates of Limnocalanus. Verh. int. Verein.
Limnol. 18: 1457–1461
Kingsland, S. E. 1985. Modeling Nature. Episodes in the History of Population Ecol-
ogy. University of Chicago Press, Chicago
Kinne, O. 1988. The scientific process — its links, functions and problems. Natur-
wissenschaften 75: 275–279
Kirchner, W. B. and P. J. Dillon. 1975. An empirical method of estimating the retention
of phosphorus in lakes. Wat. Resour. Res. 11: 181–182
Kleiber, M. 1961. The Fire of Life. Wiley, New York
Koestler, A. 1969. The Act of Creation. Macmillan, London
Kuhn, T. 1962. The Structure of Scientific Revolutions. University of Chicago Press,
Chicago
Kuhn, T. 1970. The Structure of Scientific Revolutions. 2nd edition. University of
Chicago Press, Chicago
231REFERENCES

Kuhn, T. 1977. The Essential Tension. Selected Studies in Scientific Tradition and
Change. University of Chicago Press, Chicago
Lampert, W. 1987. Feeding and assimilation in Daphnia. Memorie Ist. ital. Idrobiol. 45:
143–192
Lampert, W., W. Fleckner, H. Rai and B. E. Taylor. 1986. Phytoplankton control by
grazing zooplankton: a study on the clear-water phase. Limnol. Oceanogr. 31:
478–490
Langford, R. R. and E. G. Jermolajev. 1965. Direct effect of wind on plankton distribu-
tion. Verh. int. Verein Limnol. 16: 188–193
Larner, J. 1967. The discovery of Glycogen and Glycogen today. pp. 135–162. In:
F. Grande and M. B. Vissher (eds.) Claude Bernard and Experimental Medicine.
Schenkman Publishing Company, Inc., Cambridge
Lasenby, D. C. 1975. Development of oxygen deficits in 14 southern Ontario lakes.
Limnol. Oceanogr. 20: 993–999
Lasenby, D. C. and R. R. Langford. 1972. Growth, life history and respiration of Mysis
relicta in an arctic and temperate lake. J. Fish. Res. Bd Can. 29: 1701–1708
Lean, D. R. S. 1973. Phosphorus Compartments in Lakewater. Ph.D. Thesis, University
of Toronto, Toronto
Lehman, J. T. 1986a. Control of eutrophication in Lake Washington. pp. 301–316. In:
Ecological Knowledge and Environmental Problem Solving. National Research
Council. National Academy Press, Washington, D.C.
Lehman, J. T. 1986b. The goal of understanding in limnology. Limnol. Oceanogr. 31:
1143–1159
Liebig, J. 1840. Organic chemistry in its application to vegetable physiology and agri-
culture. pp. 12–14. In: E. J. Kormondy (ed.). Readings in Ecology. Prentice-Hall,
Englewood Cliffs, New Jersey
Likens, G. E. 1992. The Ecosystem Approach: Its Use and Abuse. In: O. Kinne (ed.).
Excellence in Ecology, Vol. 3. Ecology Institute, Oldendorf/Luhe
Lindeman, R. L. 1942. The trophic-dynamic aspect of ecology. Ecology 23: 399–418
Lonsdale, W. M. 1988. Predicting the amount of litterfall in forests of the world. Ann.
Bot. 61: 319–324
Lowry, O. H., N. J. Rosebrough, A. L. Faff and R. J. Randall. 1951. Protein measure-
ment with the folin phenol reagent. J. biol. Chem. 193: 256–265
MacArthur, R. H. 1972. Coexistence of species. pp. 253–259. In: J. Behnke (ed.). Chal-
lenging Biological Problems. Oxford University Press, Oxford
Macaulay, T. B. 1852. Lord Bacon. Longman, Brown, Green, and Longmans, London
Macdonald, C. R. and C. D. Metcalfe. 1991. Concentration and distribution of PCB
congeners in isolated Ontario lakes contaminated by atmospheric deposition. Can.
J. Fish. Aquat. Sci. 48: 371–381
Mader, S. S. 1987. Biology. Evolution, Diversity and the Environment. 2nd edition.
W. C. Brown Publishers, Dubuque, Iowa
Magee, B. 1973. Popper. Fontana-Collins, London
May, R. M. 1981. Theoretical Ecology, 2nd edition. Blackwell, Oxford
Maynard Smith, J. 1972. On Evolution. Edinburgh University Press, Edinburgh
McCarty, L. S. 1987. Relationship between toxicity and bioconcentration for some
organic chemicals. pp. 207–220. In: K. L. E. Kaiser (ed.). QSAR in Environmental
Toxicology II. D. Reidl, Dordrecht
McCauley, E., J. A. Downing and S. Watson. 1989. Sigmoid relationships between
nutrients and chlorophyll among lakes. Can. J. Fish. Aquat. Sci. 46: 1171–1175
232 REFERENCES

McCauley, E. and J. Kalff. 1981. Empirical relationships between phytoplankton and
zooplankton biomass in lakes. Can. J. Fish. Aquat. Sci. 38: 458–463
McCauley, E., W. W. Murdoch and S. Watson. 1988. Simple models and variation in
plankton densities among lakes. Am. Nat. 132: 383–403
McIntosh, R. P. 1985. The Background of Ecology: Concept and Theory. Cambridge
University Press, Cambridge
McIntosh, R. P. 1989. Citation classics of ecology. Q. Rev. Biol. 64: 31–49
McMahon, J. W. and F. H. Rigler. 1965. Feeding rates of Daphnia magna Straus in dif-
ferent foods with radioactive phosphorus. Limnol. Oceanogr. 10: 105–113
McNab, B. K. 1980. Food habits, energetics, and the population biology of mammals.
Am. Nat. 116: 106–124
McNaughton, S. J., M. Oesterheld, D. A. Frank and K. J. Willians. 1989. Ecosystem-
level patterns of primary productivity and herbivory in terrestrial habitats. Nature
341: 142–144
McQueen, D. J., J. R. Post and E. L. Mills. 1986. Trophic relationships in freshwater
pelagic ecosystems. Can. J. Fish. Aquat. Sci. 43: 1571–1581
Medawar, P. 1967. The Art of The Soluble. Methuen, London
Medawar, P. 1984. The Limits of Science. Oxford University Press, Oxford
Medawar, P. 1990. The Threat and the Glory. Harper Collins, New York
Melosh, H. J., N. M. Schneider, K. J. Zahnle and D. Latham. 1990. Ignition of global
wildfires at the Cretaceous/ Tertiary Boundary. Nature 343: 251–254
Mendelsohn, E. 1964. Heat and Life: The Development of the Theory of Animal Heat.
Harvard University Press, Cambridge
Merton, R. 1968. The Matthew effect in science. Science 159: 56–63
Miller, D. 1985. Popper Selections. Princeton University Press, Princeton
Morgan, C. L. 1923. Emergent Evolution. Williams and Norgate, London
Morin, A., C. Back, A. Chalifour, J. Boisvert and R. H. Peters. 1988a. Empirical mod-
els predicting ingestion rates of black fly larvae. Can. J. Fish. Aquat. Sci. 45:
1711–1719
Morin, A. and N. Bourassa. 1992. Modèles empiriques de la production annuelle et du
rapport P/B d’invertébrés benthiques d’eau courante. Can. J. Fish. Aquat. Sci. 49:
532–539
Morin, A., M. Constantin and R. H. Peters. 1988b. Allometric models of simuliid
growth rates and their use for estimation of production. Can. J. Fish. Aquat. Sci. 45:
315–324
Morin, A. and R. H. Peters. 1988. Effect of microhabitat features, seston quality, and
periphyton on abundance of overwintering blackfly larvae in southern Quebec.
Limnol. Oceanog. 33: 431–446
Morris, I. 1966. Is science really ‘scientific’? Science J. 1966 (Dec.): 76–80
Mosello, R., A. Calderoni and E. de Giuli. 1978. Bilancio chimico del Lago Maggiore
nel 1978. Memorie Ist. ital. Idrobiol. 39: 7–29
Naumann, E. 1930. Die Haupttypen der Gewässer in produktionsbiologischer Hinsicht.
Verh. int. Verein. Limnol. 5: 72–74
Neary, B. P. and P. J. Dillon. 1988. Effects of sulphur deposition on lake-water chem-
istry in Ontario, Canada. Nature 333: 340–343
Nicholls, K. H. and P. J. Dillon. 1978. An evaluation of phosphorus-chlorophyll-phyto-
plankton relationships for lakes. Int. Revue ges. Hydrobiol. 63: 141–154
Nixon, S. W. 1988. Physical energy inputs and the comparative ecology of lake and
marine ecosystems. Limnol. Oceanogr. 33: 1005–1025
233REFERENCES

Novaczek, I., M. S. Madhyastha, R. F. Ablett, A. Donald, G. Johnson, M. S. Nijjar, and
D. E. Sims. 1992. Depuration of domoic acid from live blue mussels (Mytilus
edulis). Can. J. Fish. Aquat. Sci. 49: 312–318
Novales-Flamarique, I., S. Griesbach, M. Parent, A. Cattaneo and R. H. Peters. 1993.
Chlorophyll and nutrient concentrations in laboratory microcosms differing in fish
foraging behaviour. Limnol. Oceanogr. 38: 290–298
Nürnberg, G. K. 1984. The prediction of internal P load in lakes with anoxic
hypolimnia. Limnol. Oceanogr. 29: 111–124
Nürnberg, G. K. and R. H. Peters. 1984. Biological availability of soluble reactive phos-
phorus in anoxic and oxic freshwaters. Can. J. Fish. Aquat. Sci. 41: 757–765
Odum, E. P. 1954. Fundamentals of Ecology, 2nd edition. W. B. Saunders Company,
Toronto
Odum, E. P. 1971. Fundamentals of Ecology, 3rd edition. W. B. Saunders Company,
Toronto
OECD. 1982. Monitoring of Inland Waters (Eutrophication Control). Synthesis Report.
OECD, Paris
O’Neil, R. V., D. L. DeAngelis, J. B. Waide and T. F. H. Allen. 1986. A Hierarchical
Concept of Ecosystems. Princeton University Press, Princeton
Orians, G. H., J. Buckley, W. Clark, M. Gilpin, C. Jordan, J. Lehman, R. May, G. Robil-
lard, D. Simberloff, W. Erckmann, D. Policansky and N. Grossblatt. 1986. Ecologi-
cal Knowledge and Environmental Problem Solving. National Academy Press,
Washington, D.C.
Oster, G. 1981. Predicting populations. Am. Zool. 21: 831–844
Ostrofsky, M. L. 1978. Modification of phosphorus retention models for use with lakes
with low areal water loading. J. Fish. Res. Bd Can. 35: 1532–1536
Pace, M. L. 1986. An empirical analysis of zooplankton community sizestructure
across lake trophic gradients. Limnol. Oceanogr. 31: 45–55
Pagel, M. D. and P. H. Harvey. 1988. Recent developments in the analysis of compara-
tive data. Q. Rev. Biol. 63: 413–440
Paine, R. T. 1977. Controlled manipulations in the marine intertidal zone, and their con-
tributions to ecological theory. pp. 245–270. In: C. E. Goulden (ed.). The Changing
scenes of Natural Sciences. Academy of Natural Sciences, Philadelphia
Patten, B. C. 1975. Systems Analysis and simulation in ecology, Volume 3. Academic
Press, New York
Patten, B. C. 1982. Environs: relativistic elementary particles for ecology. Am. Nat.
119: 179–219
Pearsall, H. W. 1932. Phytoplankton in the English lakes. II. The composition of the
phytoplankton in relation to dissolved substances. J. Ecol. 20: 241–262
Pera, M. 1980. Popper e la Scienza su Palafitte. G. Laterza e Figli Spa, Rome
Peters, R. H. 1972. Phosphorus regeneration by zooplankton. Ph.D. thesis, Univ.
Toronto, Toronto
Peters, R. H. 1976. Tautology in evolution and ecology. Am. Nat. 110: 1–12
Peters, R. H. 1977. The availability of atmospheric orthophosphate. J. Fish. Res. Bd
Can. 34: 918–924
Peters, R. H. 1980. Useful concepts for predictive ecology. pp. 215–227. In: E. Saari-
nen (ed.). Conceptual Issues in Ecology. D. Reidel, Dordrecht
Peters, R. H. 1983. The Ecological Implications of Body Size. Cambridge University
Press, Cambridge
Peters, R. H. 1984. Methods for the study of feeding, filtering and assimilation by zoo-
234 REFERENCES

plankton. pp. 336–412. In: J. A. Downing and F. H. Rigler (eds.). Secondary Pro-
ductivity in Fresh Waters. Blackwell Scientific Publications, Oxford
Peters, R. H. 1986. The role of prediction in limnology. Limnol. Oceanogr. 31:
1143–1159
Peters, R. H. 1987. Metabolism in Daphnia. Memorie Ist. ital. Idrobiol. 45: 193–243
Peters, R. H. 1989. Some pathologies in limnology. Mem. Ist. Ital. Idrobiol. 45:
175–212
Peters, R. H. 1991a. A Critique for Ecology. Cambridge University Press, Cambridge
Peters, R. H. 1991b. Lessons from the size efficiency hypothesis I. The general refuge
concept. Sel. Symp. Monogr. Union. Ital. 5: 335–361
Peters, R. H. 1992. Lessons from the size efficiency hypothesis II. The mire of com-
plexity. Hydrobiologia 235/6: 435–445
Peters, R. H. and F. H. Rigler. 1973. Phosphorus release by Daphnia. Limnol.
Oceanogr. 18: 821–839
Peters, R. H. and K. Wassenberg. 1983. The effect of body size on animal abundance.
Oecologia 60: 89–96
Pimm, S. L. and A. Redfearn. 1988. The variability of population densities. Nature 334:
613–614
Plante, C. and J. A. Downing. 1989. Production of freshwater invertebrate populations
in lakes. Can. J. Fish. Aquat. Sci. 46: 1489–1498
Platt, J. R. 1964. Strong inference. Science 146: 347–353
Polanyi, M. 1958. Personal Knowledge. University of Chicago Press, Chicago
Popper, K. R. 1934. The problem of demarcation. (1985 reprint.) pp. 118–132. In: D.
Miller (ed.). Popper Selections. Princeton University Press, Princeton
Popper, K. R. 1959. The Logic of Scientific Discovery. Harper Books, New York
Popper, K. R. and D. Miller. 1983. A proof of the impossibility of inductive probability.
Nature 302: 687–688
Porter, K. G., J. Gerritsen and J. D. Orcutt, Jr. 1982. The effect of food concentration on
swimming patterns, feeding behaviour, ingestion, assimilation and respiration by
Daphnia. Limnol. Oceanogr. 27: 935–949
Porter, K. G. and R. McDonough. 1984. The energetic cost of response to blue-green
algal filaments by cladocerans. Limnol. Oceanogr. 29: 365–369
Prairie, Y. T.-, C. M. Duarte and J. Kalff. 1989. Unifying nutrient-chlorophyll relation-
ships in lakes. Can. J. Fish. Aquat. Sci. 46: 1176–1182
Prairie, Y. T.- and J. Kalff. 1986. Effect of catchment size on phosphorus export. Wat.
Res. Bull. 22: 465–470
Pramer, D. 1985. Terminal science. BioScience 35: 141
Prepas, E. E. and F. H. Rigler. 1982. Improvements in quantifying the phosphorus con-
centration in lake water. Can. J. Fish. Aquat. Sci. Can. 39: 822–829
Price, D. J. de Solla. 1986. Little Science, Big Science....and Beyond. Columbia Uni-
versity Press, New York
Price, N. M., Andersen, L. F. and F. M. M. Morel. 1991. Iron and nitrogen nutrition of
equatorial Pacific plankton. Deep-Sea Res. 38: 1361–1378
Provasoli, L. D. E. Conklin and A. S. Agostino. 1970. Factors inducing fertility in asep-
tic Crustacea. Helgoländer wiss. Meeresunters. 20: 443–454
Pütter, A. 1909. Die Ernährung der Wassertiere und der Stoffhaushalt der Gewässer.
Gustav Fisher, Jena
Rasmussen, J. B. 1988. Littoral zoobenthic biomass in lakes, and its relationship to
physical, chemical, and trophic factors. Can. J. Fish. Aquat. Sci. 45: 1436–1447
235REFERENCES

Rasmussen, J. B. and J. Kalff. 1987. Empirical models for zoobenthic biomass in lakes.
Can. J. Fish. Aquat. Sci. 44: 990–1001
Raven, P. H. and G. B. Johnson. 1992. Biology. 3rd edition. Times Mirror/Mosby Col-
lege Publishing, St. Louis, Missouri
Rawson, D. S. 1955. Morphometry as a dominant factor in the productivity of large
lakes. Verh. int. Verein. Limnol. 12: 164–175
Reckhow, K. H. and S. C. Chapra. 1983. Engineering Approaches to Lake Manage-
ment. Volume 1. Data Analysis and Empirical Modelling. Butterworth, Woburn,
Massachusetts
Reckhow, K. H. and J. T. Simpson. 1980. A procedure using modelling and error anal-
ysis for the prediction of lake phosphorus concentration from land use information.
Can. J. Fish. Aquat. Sci. 37: 1439–1448
Remmert, H. 1980. Ecology. Springer-Verlag, Berlin
Ricker, W. E. 1984. Computation and use of central trend lines. Can. J. Zool. 62:
1897–1905
Rigler, F. H. 1956. A tracer study of the phosphorus cycle in lake water. Ecology 37:
550–562
Rigler, F. H. 1964. The phosphorus fractions and the turnover time of inorganic phos-
phorus in different types of lakes. Limnol. Oceanogr. 9: 511–518
Rigler, F. H. 1966. Radiobiological analysis of inorganic phosphorus in lakewater.
Verh. int. Verein. Limnol. 16: 465–470
Rigler, F. H. 1971. Feeding rates. pp. 228–255. In: W. T. Edmondson and G. G. Win-
berg (eds.). Secondary Productivity of Fresh Waters. Blackwell Scientific Publica-
tions, Oxford
Rigler, F. H. 1974. Appendix. Phosphorus cycling in lakes. pp. 263–273. In: F. Ruttner
(ed.). Fundamentals of Limnology. University of Toronto Press, Toronto
Rigler, F. H. 1975a. Nutrient kinetics and the new typology. Verh. int. Verein. Limnol.
19: 197–210
Rigler, F. H. 1975b. The concept of energy flow and nutrient flow between trophic
levels. pp. 15–26. In: W. H. van Dobben and R. H. Lowe-McConnell (eds.). Unify-
ing Concepts in Ecology. Dr. W. Junk, The Hague
Rigler, F. H. 1976. Review of Patten, B. C. (ed.) 1975. Systems analysis and simulation
in ecology, Volume 3. Academic Press, New York. Limnol. Oceanogr. 21: 481–483
Rigler, F. H. 1978. Limnology in the high Arctic: a case study of Char Lake. Verh. int.
Verein. Limnol. 20: 127–140
Rigler, F. H. 1982a. The relation between fisheries management and limnology. Trans.
Am. Fish. Soc. 111: 121–132
Rigler, F. H. 1982b. Recognition of the possible: An advantage of empiricism in ecol-
ogy. Can. J. Fish. Aquat. Sci. 39: 1323–1331
Rigler, F. H. and J. A. Downing. 1984. The calculation of secondary productivity.
pp. 19–46. In: J. A. Downing and F. H. Rigler (eds.). A manual on methods for the
assessment of secondary productivity in fresh waters. Blackwell Scientific Publica-
tions, London
Rigler, F. H. and R. R. Langford. 1967. Congeneric occurrences of species of in south-
ern Ontario lakes. Can. J. Zool. 45: 81–90
Rigler, F. H., M. E. MacCallum and J. C. Roff. 1974. Production of zooplankton in Char
Lake. J. Fish. Res. Bd Can. 31: 637–646
Robinson, W. R., R. H. Peters and J. Zimmerman. 1983. The effects of body size and
temperature on metabolic rate of organisms. Can. J. Zool. 61: 281–288
236 REFERENCES

Roe, A. 1953. The Making of a Scientist. Dodd-Mead, New York
Rose, L. 1972. The censorship of Velikovsky’s interdisciplinary synthesis. Pensée 2 (2):
29–31
Rowan, D. J., J. Kalff and J. B. Rasmussen. 1992. Estimating the mud deposition
boundary depth in lakes from wave theory. Can. J. Fish. Aquat. Sci. 49: 2490–2497
Ruse, M. 1973. The Philosophy of Biology. Hutchinson University Library, London
Ruse, M. 1982. Darwinism Defended: A Guide to the Evolution Controversies. Addi-
son-Wesley, Reading, Massachusetts
Russell, B. 1931. The scientific outlook. W. W. Norton, New York
Ryder, R. A. 1965. A method for estimating the potential fish production of north tem-
perate lakes. Trans. Am. Fish. Soc. 94: 214–218
Ryder, R. A. 1982. The morphoedaphic index - use, abuse and fundamental concepts.
Trans. Am. Fish. Soc. 111: 154–64
Sakamoto, M. 1966. Primary production by phytoplankton community in some
Japanese lakes and its dependence on lake depth. Arch. Hydrobiol. 62: 1–28
Sandercock, G. A. 1967. A study of selected mechanisms for the coexistence of Diap-
tomus spp. in Clarke lake, Ontario. Limnol. Oceanogr. 12: 97–112
Sas, H. 1989. Lake Restoration by Reduction of Nutrient Loading. Springer-Verlag,
Berlin
Sattler, R. 1986. Biophilosophy: Analytic and Holistic Perspectives. Springer-Verlag,
Berlin
Scheider, W. A. 1978. Applicability of phosphorus budget models to small precambrian
lakes, Algonquin Park, Ontario. Can. J. Fish. Aquat. Sci. 35: 300–304
Schindler, D. W. 1971. Carbon, nitrogen and phosphorus and the eutrophication of
freshwater lakes. J. Phycol. 7: 321–322
Schindler, D. W. S. 1974. Eutrophication and recovery in experimental lakes: implica-
tions for lake management. Science 184: 897–899
Schindler, D. W. 1978. Evolution of phosphorus limitation in lakes. Science 196:
260–262
Schindler, D. W. and E. J. Fee. 1974. Experimental Lakes area: whole-lake experiments
in eutrophication. Can. J. Fish. Aquat. Sci. 31: 937–953
Schmidt, G. W. 1968. Zur Ausnutzung des Nahrungsstickstoffs durch Daphnia magna
Straus. Arch. Hydrobiol. 65: 142–186
Schoener, T. W. 1972. Mathematical ecology and its place among the sciences. I. The
biological domain. Science 178: 389–394
Schoener, T. W. 1985. Are lizard population sizes unusually consistent through time.
Am. Nat. 126: 633–641
Schrader-Frechette, K. S. and E. D. McCoy. 1993. Method in Ecology. Cambridge Uni-
versity Press, Cambridge
Science Council of Canada. 1988. Water 2020: Sustainable Use for Water in the 21st
Century. The Publications Office, Ottawa
Seim, E. and B.-E. Saether. 1983. On rethinking allometry: Which regression model to
use? J. theor. Biol. 104: 161–168
Seip, K. L. and H. Ibrekk. 1988. Regression equations for lake management — how far
do they go. Verh. int. Verein. Limnol. 23: 778–785
Shapiro, J. 1978. The need for more biology in lake restoration. USEPA National Con-
ference on Lake Restoration. Minneapolis, Minnesota
Shapiro, J. and D. I. Wright. 1984. Lake restoration by biomanipulation: Round Lake,
Minnesota, the first two years. Freshwat. Biol. 14: 371–83
237REFERENCES

Shaw, G. B. 1921. Back to Methuselah. A Metabiological Pentateuch. Constable, Lon-
don
Shaw, G. B. 1931. Man and Superman. A comedy and a philosophy. Constable, London
Shelford, V. E. 1911. Physiological animal geography. J. Morph. 22: 551–618
Slobodkin, L. B. 1968. Towards a predictive theory of evolution. pp. 317–340. In:
R. Lewontin (ed.). Population biology and evolution. Syracuse University Press,
Syracuse, New York
Smayda, T. J. 1974. Bioassay of the growth potential of the surface water of lower Nar-
ragansett Bay over an annual cycle using the diatom Thalassiosira pseudonana
(oceanic clone 13–1). Limnol. Oceanogr. 19: 889–901
Smith, R. J. 1980. Rethinking allometry. J. theor. Biol. 87: 87–111
Smith, V. H. 1979. Nutrient dependence of primary productivity in lakes. Limnol.
Oceanogr. 24: 1051–1064
Smith, V. H. 1982. The nitrogen and phosphorus dependence of algal biomass in lakes:
an empirical and theoretical analysis. Limnol. Oceanogr. 27: 1101–11
Smith, V. H., F. H. Rigler, O. Choulik, M. Diamond, S. Griesbach and D. Skraba. 1984.
Effects of phosphorus fertilization on phytoplankton biomass and phosphorus reten-
tion in subarctic Quebec lakes. Verh. int. Verein. Limnol. 22: 376–382
Snow, C. P. 1963. The Two Cultures: and a Second Look. Mentor Books, New York
Sommer, U., Z. M. Gliwicz, W. Lampert and A. Duncan. 1986. The Plankton Ecology
Group (PEG) model of seasonal succession of planktonic events in fresh waters.
Arch. Hydrobiol. 106: 433–471
Southwood, T. R. E. 1988. Tactics, strategies and templets. Oikos 52: 3–17
Stephens, D. W. and J. R. Krebs. 1986. Foraging Theory. Princeton University Press,
Princeton
Stove, D. 1972. The scientific mafia. Pensée 2(2) 6–8, 49
Strasˇkraba, M. 1980. The effects of physical variables on freshwater production: Anal-
yses based on models. pp. 13–84. In: E. D. LeCren and R. H. Lowe-McConnell
(eds.). The Functioning of Freshwater Ecosystems. Cambridge University Press,
Cambridge
Tansley, A. G. 1935. The use and abuse of vegetational concepts and terms. Ecology 16:
284–307
Tarapchak, S. J. and L. R. Herche. 1988. Orthophosphate concentrations in lake water:
analysis of Rigler’s radiobioassay method. Can. J. Fish. Aquat. Sci. 45: 2230–2237
Taubes, G. 1993. Measure for measure in Science. Science 260: 884–886
Taylor, W. D., J. H. Carey, D. R. S. Lean and D. J. McQueen. 1991. Organochlorine
concentrations in the plankton of lakes in southern Ontario and their relationship to
plankton biomass. Can. J. Fish. Aquat. Sci. 48: 1960–1966
Taylor, W. D. and D. R. S. Lean. 1991. Phosphorus pool sizes and fluxes in the epil-
imnion of a mesotrophic lake. Can. J. Fish. Aquat. Sci. 48: 1293–1301
Tessier, A. J. and C. E. Goulden 1987. Cladoceran juvenile growth. Limnol. Oceanogr.
32: 680–686
Thienemann, A. 1926. Der Nahrungkreislauf im Wasser. Verh. dt. zool. Ges. 31: 29–79
Thomas, E. A. 1969. The process of eutrophication in central european lakes.
pp. 29–49. In: Eutrophication: Causes, Consequences, Correctives. Proceedings of
a Symposium. National Academy of Sciences, Washington, D.C.
Tucker, A. 1957. The relation of phytoplankton periodicity to the nature of the physico-
chemical environment with special reference to phosphorus. Am. Midl. Nat. 57:
300–370
238 REFERENCES

Turner, J. T., P. A. Tester and J. R. Strickler. 1993. Zooplankton feeding ecology: A cin-
ematographic study of animal-to-animal variability in the feeding behaviour of
Calanus finmarchicus. Limnol. Oceanogr. 38: 255–264
van Straten, G. and K. J. Keesman 1991. Uncertainty propagation and speculation in
projective forecasts of environmental change: a lake-eutrophication example. J.
Forcast. 10: 163–190
Velikovsky, I. 1950. Worlds in Collision. MacMillan, New York
Velikovsky, I. 1955. Earth in Upheaval. Doubleday, New York
Verschueren, K. 1983. Handbook of Environmental Data on Organic Chemicals. Van
Nostrand Reinhold, New York
Vollenweider, R. A. 1968. Scientific Fundamentals of Eutrophication of Lakes and
Flowing Waters with Special Reference to Phosphorus and Nitrogen. OECD, Paris.
OECD/DAS/SCI/68.27
Vollenweider, R. A. 1969. Möglichkeiten und Grenzen elementarer Modelle der Stoff-
bilanz von Seen. Arch. Hydrobiol. 66: 1–36
von Bertalanffy, L. 1950. The theory of open systems in physics and biology. Science
111: 23–29
von Bertalanffy, L. 1952. Problems of life. C. A. Watts, London
Webster, K. E. and R. H. Peters. 1978. Some size dependent inhibitions of larger clado-
ceran filterers in filamentous suspensions. Limnol. Oceanogr. 23: 1138–1145
Weisz, P. B. and R. N. Keogh. 1982. The Science of Biology. 5th edition. McGraw-Hill
Book Co., New York
Welch, H. E. 1974. Metabolic rates of arctic lakes. Limnol. Oceanogr. 19: 65–73
Welch, H. E. 1976. Ecology of Chironomidae (Diptera) in a polar lake. J. Fish. Res. Bd
Can. 33: 227–247
Welch, H. E. and J. Kalff. 1974. Benthic photosynthesis and respiration in Char Lake.
J. Fish. Res. Bd Can. 31: 609–620
Wetzel, R. G. 1991. Limnological education — reply to the comment by Kalff. Limnol.
Oceanogr. 36: 1502
Whitehead, A. N. 1925. Science and the Modern World. (Anthology edition, 1953.)
pp. 363–466. In: F. S. C. Northrop and M. W. Gross (eds.). Alfred North Whitehead:
An Anthology. Cambridge University Press, Cambridge
Wimsatt, W. C. 1980. Reductionistic research strategies and their biases in the units of
selection controversy. pp. 155–202. In: E. Saarinen (ed.). Conceptual Issues in
Ecology. D. Reidel, Dordrecht
Zar, J. H. 1968. Calculation and miscalculation of the allometric equation as a model in
biological data. BioScience. 18: 1118–1120
Zuckerman, H. A. 1977. Scientific Elite: Nobel Laureates in the United States. Free
Press, New York
239REFERENCES

About the Author and the Book
Professor Robert H. Peters is the winner of the
ECOLOGY INSTITUTE PRIZE 1991 in limnetic
ecology. Born in 1946 in Toronto, Canada, he ob-
tained his Ph.D. in 1972 from the University of
Toronto under the supervision of Frank H. Rigler, a
major figure in limnology, who died much too
early, leaving behind a host of important work and
thought. In his EE book, Peters presents highlights
of Rigler’s unpublished notes and ideas and com-
bines them masterfully with his own accomplish-
ments and expertise.
Chaired by Professor Jürgen Overbeck (Max-
Planck-Institut für Limnologie, Plön, Germany)
the ECI Jury selected Rob Peters for his work on
phosphorus cycling in lakes, which provides exam-
ples of excellent research and illuminates impor-
tant insights into the measurement and availability
of phosphorus in aquatic systems.
This book reaches far into the realms of science history, philosophy and methodology,
the significance of science for society, and the research and teaching in universities. It
documents that ecologists have collected impressive amounts of observations and facts,
but that they have failed to sufficiently identify and formulate theories which go beyond
the facts — theories that can be tested and that can predict.
About the Ecology Institute (ECI)*
The international ECI is a not-for-profit organization of research ecologists. Director and
scientific staff — 52 marine, terrestrial and limnetic ecologists of outstanding profes-
sional reputation — strive to honor excellence in ecological research; to further the
exchange among marine, terrestrial and limnetic ecologists; to promote advancement in
environmental sciences; and to bridge the gap between ecological science and its appli-
cation for the benefit of nature and society.
In order to approach these goals the ECI annually sets out two international prizes, the
ECI and IRPE (International Recognition of Professional Excellence) Prize, and it sup-
ports — via the Otto Kinne Foundation (OKF) — promising young environmental scien-
tists in Eastern European countries by providing financial assistance for professional
travel, scientific equipment or published information. Each ECI Prize Laureate is re-
quested to author a book taking into account ECI’s aims. The book is published in the
series “Excellence in Ecology” and made available worldwide at cost price; a consider-
able number of books are donated to libraries in Third-World countries. In this way
leading ecologists are offered the possibility of disseminating their personal views on
current ecological issues and of serving the general public who depend acutely on defini-
tive ecological knowledge for planning our present and future.
* Nordbünte 23, D-21385 Oldendorf/Luhe, Germany
Tel. (+49) (0) 4132 7127; Fax (+49) (0) 4132 8883; E-mail 100327[email protected]
Robert H. Peters
